Ιατρικός εμπειρογνώμονας του άρθρου

Νέες δημοσιεύσεις
Λύσσα στα παιδιά
Τελευταία επισκόπηση: 04.07.2025

Όλα τα περιεχόμενα του iLive ελέγχονται ιατρικά ή ελέγχονται για να διασφαλιστεί η όσο το δυνατόν ακριβέστερη ακρίβεια.
Έχουμε αυστηρές κατευθυντήριες γραμμές προμήθειας και συνδέουμε μόνο με αξιόπιστους δικτυακούς τόπους πολυμέσων, ακαδημαϊκά ερευνητικά ιδρύματα και, όπου είναι δυνατόν, ιατρικά επισκοπικά μελέτες. Σημειώστε ότι οι αριθμοί στις παρενθέσεις ([1], [2], κλπ.) Είναι σύνδεσμοι με τις οποίες μπορείτε να κάνετε κλικ σε αυτές τις μελέτες.
Εάν πιστεύετε ότι κάποιο από το περιεχόμενό μας είναι ανακριβές, παρωχημένο ή αμφισβητήσιμο, παρακαλώ επιλέξτε το και πατήστε Ctrl + Enter.
Η λύσσα, ή υδροφοβία, είναι μια οξεία ιογενής ασθένεια που μεταδίδεται μέσω του δαγκώματος ενός μολυσμένου ζώου, με βλάβη στο νευρικό σύστημα και την ανάπτυξη σοβαρής εγκεφαλίτιδας με θανατηφόρο έκβαση.
Επιδημιολογία
Μια μάστιγα για τη δημόσια υγεία από την αρχαιότητα, ο ιός της λύσσας προκαλεί σήμερα περίπου 59.000 θανάτους σε ανθρώπους κάθε χρόνο, σχεδόν όλοι εκ των οποίων μεταδίδονται από δαγκώματα σκύλων. Αυτό έχει σημαντικό οικονομικό αντίκτυπο στις αναπτυσσόμενες χώρες, ιδιαίτερα στην Αφρική και την Ασία, οι οποίες μπορούν να υποστούν τις λιγότερες τέτοιες απώλειες. Ωστόσο, παρά το ποσοστό θνησιμότητας που φτάνει σχεδόν το 100%, η λύσσα των σκύλων είναι μια ασθένεια που μπορεί να προληφθεί πλήρως και ιστορικά παραδείγματα εξάλειψης της λύσσας στους σκύλους στον ανεπτυγμένο κόσμο το επιβεβαιώνουν αυτό. [ 1 ]
Αιτίες λύσσα
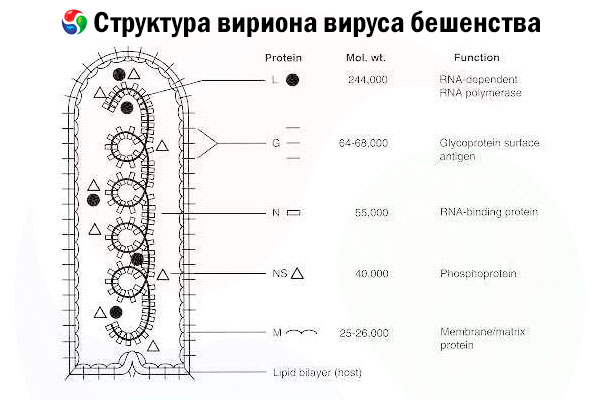
Ο αιτιολογικός παράγοντας είναι ο ιός της λύσσας (RV), ένας ιός αρνητικής αλυσίδας RNA της οικογένειας των ραβδοϊών, μεγέθους περίπου 60 nm × 180 nm.
Αποτελείται από έναν εσωτερικό πρωτεϊνικό πυρήνα, ή νουκλεοκαψίδιο, που περιέχει νουκλεϊκό οξύ, και μια εξωτερική μεμβράνη, μια διπλοστιβάδα που περιέχει λιπίδια και καλύπτεται με διαμεμβρανικές αιχμές γλυκοπρωτεϊνών. Έχει μια σχετικά απλή αρθρωτή δομή γονιδιώματος και κωδικοποιεί πέντε δομικές πρωτεΐνες:
- RNA-εξαρτώμενη RNA πολυμεράση (L),
- νουκλεοπρωτεΐνη (Ν),
- φωσφορυλιωμένη πρωτεΐνη (P),
- πρωτεΐνη μήτρας (Μ) και
- γλυκοπρωτεΐνη εξωτερικής επιφάνειας (G).
Οι πρωτεΐνες N, P και L μαζί με το γονιδιωματικό RNA σχηματίζουν το σύμπλεγμα ριβονουκλεοπρωτεϊνών. Το G είναι το μόνο αντιγόνο του ιού των ιών των ιών των ιών που είναι ικανό να προκαλέσει την παραγωγή αντισωμάτων εξουδετέρωσης του ιού των ιών των ιών, τα οποία είναι οι κύριοι ανοσοποιητικοί παράγοντες κατά της θανατηφόρας λοίμωξης από ιό των ιών των ιών. Από την άλλη πλευρά, το σύμπλεγμα ριβονουκλεοπρωτεϊνών έχει αποδειχθεί ότι είναι το κύριο αντιγόνο του ιού των ιών των ιών που είναι ικανό να προκαλέσει την παραγωγή CD4+ Τ κυττάρων, τα οποία μπορούν να ενισχύσουν την παραγωγή αντισωμάτων εξουδετέρωσης του ιού των ιών των ιών μέσω της ενδοδομικής αναγνώρισης αντιγόνου.[ 2 ] Το σύμπλεγμα ριβονουκλεοπρωτεϊνών μπορεί να παίζει σημαντικό ρόλο στην καθιέρωση της ανοσολογικής μνήμης και της μακροχρόνιας ανοσίας.[ 3 ]
 [
[ Ταξινόμηση και τύποι αντιγόνων
Το γένος Lyssavirus περιλαμβάνει τον ιό της λύσσας και αντιγονικά και γενετικά συγγενείς ιούς λύσσας: τους ιούς νυχτερίδας Lagos, Mokola και Duvenhage, καθώς και δύο πιθανούς υποτύπους των ιών λύσσας των ευρωπαϊκών νυχτερίδων. Μελέτες διασταυρούμενης προστασίας δείχνουν ότι τα ζώα που έχουν ανοσοποιηθεί με παραδοσιακά εμβόλια λύσσας ενδέχεται να μην προστατεύονται πλήρως όταν εκτίθενται σε άλλους ιούς λύσσας.
Οι ιοί λύσσας μπορούν να ταξινομηθούν ως σταθεροί (προσαρμοσμένοι μέσω διέλευσης σε ζώα ή κυτταροκαλλιέργειας) ή οδικοί (άγριου τύπου). Η χρήση μονοκλωνικών αντισωμάτων και γενετικής αλληλούχισης για τη διαφοροποίηση των ιών λύσσας της οδού έχει βοηθήσει στην αναγνώριση ιικών παραλλαγών που προέρχονται από μεγάλες δεξαμενές ξενιστών παγκοσμίως και στην υπόδειξη πιθανών πηγών ανθρώπινης έκθεσης όταν δεν υπήρχε ιστορικό οριστικού δαγκώματος ζώου στην περίπτωση ενός ασθενούς.[ 8 ]
Παθογένεση
Η κύρια δεξαμενή και πηγή μόλυνσης μεταξύ των άγριων ζώων είναι οι λύκοι, οι αλεπούδες, τα τσακάλια, οι νυχτερίδες, και μεταξύ των κατοικίδιων ζώων - σκύλοι και γάτες, σπάνια - άλογα, βοοειδή, χοίροι, αρουραίοι κ.λπ. Η μετάδοση της λοίμωξης από άτομο σε άτομο, αν και πιθανή, είναι εξαιρετικά σπάνια. Πρόκειται για μια τυπική ζωονόσο. Οι άνθρωποι μολύνονται με λύσσα κυρίως από σκύλους.
Αφού ένας άνθρωπος δαγκωθεί από ένα άρρωστο ζώο, ο ιός πολλαπλασιάζεται στον μυϊκό ιστό στο σημείο του δαγκώματος και στη συνέχεια, έχοντας φτάσει στα άκρα των αισθητήριων περιφερειακών νεύρων, εξαπλώνεται κεντρομόλος, φτάνοντας στους κινητικούς νευρώνες. Ο χρόνος που χρειάζεται ο ιός για να κινηθεί και να επηρεαστεί ο εγκέφαλος εξαρτάται από το σημείο του δαγκώματος. Σε περίπτωση σοβαρών δαγκωμάτων στο κεφάλι και το πρόσωπο, ο ιός μπορεί να φτάσει στο κεντρικό νευρικό σύστημα σε 15-20 ημέρες, και σε περίπτωση μικρής βλάβης στο δέρμα του κορμού και των άκρων και, κατά συνέπεια, μιας μικρής δόσης του παθογόνου, η διαδικασία μεταφοράς του ιού στο κεντρικό νευρικό σύστημα μπορεί να καθυστερήσει για αρκετούς μήνες ή και έως και 1-1,5 χρόνια. Αφού φτάσει στο κεντρικό νευρικό σύστημα, ο ιός σταθεροποιείται στους ιστούς του εγκεφάλου και του νωτιαίου μυελού, κυρίως στους νευρώνες του προμήκη μυελού, του κέρατος του Άμμωνα και της βάσης του εγκεφάλου. Στον νωτιαίο μυελό, τα οπίσθια κέρατα επηρεάζονται περισσότερο. Από το κεντρικό νευρικό σύστημα, ο ιός φυγοκεντρικά κατά μήκος των νευρικών κορμών φτάνει στους σιελογόνους αδένες, όπου πολλαπλασιάζεται και απεκκρίνεται με σάλιο.
Έννοιες στην παθογένεση της λύσσας
Ο RV έχει ένα ευρύ φάσμα ξενιστών και μπορεί να μολύνει σχεδόν όλα τα θηλαστικά. Παρόλο που έχουν αναφερθεί αρκετές οδοί μετάδοσης του RV, η φυσική μόλυνση συμβαίνει συνήθως μέσω δαγκώματος. Εκτός από τα δαγκώματα, η κατανάλωση πτωμάτων μολυσμένων με RV μπορεί να προωθήσει τη μόλυνση από τον ιό της λύσσας στις αρκτικές αλεπούδες, και η επαφή του RV με τους βλεννογόνους έχει βρεθεί ότι είναι μια άλλη πιθανή οδός μετάδοσης.[ 9 ] Σε ορισμένες ασυνήθιστες περιπτώσεις, όπως η τυχαία απελευθέρωση RV ως αεροζόλ σε εργαστήριο ή RV ως αεροζόλ σε σπηλιές που κατοικούνται από μεγάλο αριθμό νυχτερίδων,[ 10 ] μπορεί να συμβεί μετάδοση μέσω αεροζόλ.
Δεν είναι ακόμη σαφές εάν τα στελέχη του ιού του δρόμου (street RV) και τα στελέχη του προσαρμοσμένα σε ποντίκια ή σε ιστοκαλλιέργειες αναπαράγονται στο σημείο του εμβολιασμού πριν εισέλθουν στο ΚΝΣ. Ενώ η πειραματική ενδομυϊκή μόλυνση νεαρών χάμστερ ή ρακούν με τον ιό του δρόμου αποκάλυψε την αναπαραγωγή του RV σε γραμμωτά μυϊκά κύτταρα πριν ο ιός εισβάλει στους νευράξονες των κινητικών νευρώνων διαμέσου των νευρομυϊκών συνδέσεων,[ 11 ],[ 12 ] η ενδομυϊκή μόλυνση ποντικών με προσαρμοσμένο σε ποντίκια RV CVS-24 έδειξε ότι ο RV μεταναστεύει απευθείας στο ΚΝΣ χωρίς προηγούμενη αναπαραγωγή στο σημείο του εμβολιασμού.[ 13 ] Μόλις βρεθεί στα άκρα των μη μυελινωμένων αξόνων, ο RV μεταφέρεται ανάδρομα στο κυτταρικό σώμα.
Πρόσφατα ευρήματα υποδηλώνουν ότι η μεταφορά μέσω αξονικών κυστιδίων μπορεί να αντιπροσωπεύει μια βασική στρατηγική για την μετακίνηση ιικών σωματιδίων σε μεγάλες αποστάσεις στους άξονες.[ 14 ] Έχει εκτιμηθεί ότι ο ιός του ιού της πνευμονικής αρτηρίας (RV) μεταναστεύει εντός των αξόνων με ρυθμό 3 mm/h.[ 15 ] Η λοίμωξη στη συνέχεια εξαπλώνεται μέσω μιας αλυσίδας νευρώνων που συνδέονται με συναπτικές συνδέσεις. Ωστόσο, ο ακριβής μηχανισμός που προάγει τη διασυναπτική εξάπλωση είναι ακόμη άγνωστος. Μετά τη μόλυνση του εγκεφάλου, ο ιός εξαπλώνεται φυγοκεντρικά στο περιφερικό και αυτόνομο νευρικό σύστημα σε πολλά περιφερειακά όργανα.[ 16 ] Στο τελευταίο στάδιο του κύκλου της λοίμωξης, ο ιός της πνευμονικής αρτηρίας μεταναστεύει στους σιελογόνους αδένες. Μετά την αντιγραφή του σε βλεννογόνα ακινικά κύτταρα, απελευθερώνεται στο σάλιο και είναι έτοιμος για μετάδοση στον επόμενο ξενιστή.[ 17 ]
Όσον αφορά την παθολογία που προκαλείται από τον ιό της λύσσας, ο αποπτωτικός κυτταρικός θάνατος έχει προταθεί ως πιθανός παθογενετικός μηχανισμός σε πειραματικά μοντέλα λύσσας ποντικών μολυσμένων με ένα σταθερό στέλεχος του ιού της λύσσας.[ 18 ] Ένας παθογενετικός μηχανισμός που μπορεί να συμβάλλει στη βαθιά δυσλειτουργία του ΚΝΣ που χαρακτηρίζει τη λύσσα μπορεί να είναι η μειωμένη νευρωνική λειτουργία. Η γονιδιακή έκφραση έχει αποδειχθεί ότι είναι σημαντικά μειωμένη σε νευρώνες μολυσμένους με ιό του ιού της λύσσας, με αποτέλεσμα μια γενική καταστολή της πρωτεϊνικής σύνθεσης,[ 19 ] και αρκετές μελέτες έχουν δείξει μειωμένη νευροδιαβίβαση μετά από μόλυνση από ιό του ιού της λύσσας. Ο Jiang απέδειξε ότι η σύνδεση ενός ανταγωνιστή του υποδοχέα της ακετυλοχολίνης σε μολυσμένα ομογενοποιήματα εγκεφάλου αρουραίου ήταν μειωμένη σε σύγκριση με τους μάρτυρες.[ 20 ] Παρατηρήθηκε επίσης μειωμένη απελευθέρωση και σύνδεση της σεροτονίνης, ενός νευροδιαβιβαστή που εμπλέκεται στον έλεγχο του κύκλου ύπνου, της αντίληψης του πόνου και της συμπεριφοράς, σε εγκέφαλο αρουραίου μολυσμένο με ιό του ιού της λύσσας.[ 21 ],[ 22 ] Εκτός από την επίδραση στη νευροδιαβίβαση, η λοίμωξη της δεξιάς κοιλίας μπορεί επίσης να επηρεάσει τα ιοντικά κανάλια. Τα μολυσμένα κύτταρα νευροβλαστώματος ποντικού εμφανίζουν μειωμένη λειτουργική έκφραση διαύλων νατρίου που ελέγχονται από τάση, γεγονός που μπορεί να αποτρέψει τα δυναμικά δράσης και τελικά να οδηγήσει σε λειτουργική βλάβη. [ 23 ]
Εκτός από την απουσία σοβαρών παθολογικών αλλοιώσεων στο ΚΝΣ, οι περισσότερες περιπτώσεις ανθρώπινης λύσσας δεν προκαλούν ανοσολογική απόκριση 7 έως 10 ημέρες μετά την έναρξη των κλινικών συμπτωμάτων. Αυτές οι βαθιές διαφορές μεταξύ της παθογένεσης της λύσσας και αυτής των περισσότερων άλλων ιογενών ή βακτηριακών λοιμώξεων του ΚΝΣ υποστηρίζονται περαιτέρω από το γεγονός ότι η ανοσοκαταστολή είναι είτε αναποτελεσματική είτε επιζήμια για την έκβαση της λύσσας.[24 ] Το χαμηλό επίπεδο ανοσολογικής απόκρισης που παρατηρείται συχνά σε θύματα λύσσας είναι αινιγματικό, επειδή δεν μπορεί να εξηγηθεί από την κακή ανοσογονικότητα των αντιγόνων του ιού RV. Στην πραγματικότητα, η πρωτεΐνη G του ιού RV και η πρωτεΐνη νουκλεοκαψιδίου είναι ισχυρά αντιγόνα Β- και Τ-κυττάρων όταν χορηγούνται παρεντερικά. [ 25 ] Μια πιθανή εξήγηση για τον χαμηλό βαθμό ανοσολογικής απόκρισης έναντι του ιού RV σε ανθρώπους ή ζώα με λύσσα μπορεί να είναι ότι η μόλυνση του ΚΝΣ από ιό RV προκαλεί ανοσοκαταστολή, [ 26 ] και έχει προταθεί ότι ο ιός RV χρησιμοποιεί μια ανατρεπτική στρατηγική που περιλαμβάνει την πρόληψη της απόπτωσης και την καταστροφή των εισβάλλοντων Τ κυττάρων. [ 27 ]
Τα εξασθενημένα στελέχη RV που έχουν προσαρμοστεί σε μη νευρωνικά κύτταρα διαφέρουν σημαντικά από τα παθογόνα στελέχη RV του δρόμου ως προς τη νευροδιηθητικότητά τους, η οποία αναφέρεται στην ικανότητά τους να εισβάλλουν στο ΚΝΣ από περιφερειακές θέσεις. Από αυτή την άποψη, τα στελέχη RV που έχουν προσαρμοστεί σε ιστοκαλλιέργειες είτε δεν έχουν είτε έχουν μόνο περιορισμένη ικανότητα να εισβάλλουν στο ΚΝΣ από περιφερειακές θέσεις, ενώ τα στελέχη RV του δρόμου ή τα στελέχη RV που έχουν προσαρμοστεί σε ποντίκια, όπως το CVS-24, είναι ιδιαίτερα διηθητικά.[ 28 ] Βασικοί παράγοντες που εμπλέκονται στη νευροδιείσδυση του RV περιλαμβάνουν την ιική πρόσληψη, την αξονική μεταφορά, τη διασυναπτική εξάπλωση και τον ρυθμό αντιγραφής του ιού.
Μέχρι πρόσφατα, οι γνώσεις μας για την παθογένεση του ιού της λύσσας ήταν περιορισμένες και βασίζονταν κυρίως σε περιγραφικές μελέτες στελεχών του ιού της λύσσας που διαδίδονται στο δρόμο ή σε πειραματικές λοιμώξεις με εξασθενημένα στελέχη που έχουν προσαρμοστεί στο εργαστήριο. Η έλευση της τεχνολογίας της αντίστροφης γενετικής μας επέτρεψε να εντοπίσουμε τα ιικά στοιχεία που καθορίζουν τον παθογενετικό φαινότυπο του ιού της λύσσας και να κατανοήσουμε καλύτερα τους μηχανισμούς που εμπλέκονται στην παθογένεση της λύσσας.
Ταυτοποίηση ιικών στοιχείων που ελέγχουν την απόκτηση, τη διασπορά και την αναπαραγωγή του ιού της λύσσας
- Ιικά στοιχεία που εμπλέκονται στη σύλληψη του ιού
Η λοίμωξη από ιό RV ξεκινά με την προσκόλληση του ιού σε έναν πιθανολογούμενο κυτταρικό υποδοχέα. Παρόλο που πολλά μόρια στην επιφάνεια της μεμβράνης έχουν προταθεί ως υποδοχείς RV, συμπεριλαμβανομένου του νικοτινικού υποδοχέα ακετυλοχολίνης,[ 29 ] του μορίου προσκόλλησης νευρικών κυττάρων[ 30 ] και του υποδοχέα νευροτροφίνης p75 NTR χαμηλής συγγένειας,[ 31 ] εξακολουθεί να μην είναι σαφές εάν αυτά τα μόρια παίζουν πράγματι ρόλο στον κύκλο ζωής του ιού της λύσσας. Σε αυτό το πλαίσιο, πρόσφατα αποδείχθηκε ότι η αλληλεπίδραση RV G–p75 NTR δεν απαιτείται για τη μόλυνση πρωτογενών νευρώνων από RV.[ 32 ] Μετά τη σύνδεση με τον υποδοχέα, ο RV εσωτερικεύεται μέσω προσροφητικής ή ενδοκυττάρωσης που προκαλείται από υποδοχέα.[ 33 ] Το περιβάλλον χαμηλού pH εντός του ενδοσωμικού διαμερίσματος προκαλεί στη συνέχεια αλλαγές διαμόρφωσης στο RV G που πυροδοτούν τη σύντηξη της ιικής μεμβράνης με την ενδοσωμική μεμβράνη, απελευθερώνοντας έτσι το RNP στο κυτταρόπλασμα.[ 34 ] Για τους ιούς, το RV G παίζει κρίσιμο ρόλο στην ιική πρόσληψη, πιθανότατα μέσω αλληλεπιδράσεων με πιθανούς κυτταρικούς υποδοχείς που διευκολύνουν την ταχεία πρόσληψη. Από αυτή την άποψη, έχει αποδειχθεί ότι η παθογένεια των στελεχών RV προσαρμοσμένων σε καλλιέργεια ιστών (π.χ., ERA, HEP και CVS-11) συσχετίζεται με την παρουσία ενός καθοριστικού παράγοντα που βρίσκεται στην αντιγονική θέση III της πρωτεΐνης G. [ 35 ] Μια μετάλλαξη Arg → Gln στη θέση 333 σε αυτήν την αντιγονική θέση της πρωτεΐνης ERA G είχε ως αποτέλεσμα μια επταπλάσια καθυστέρηση στην εσωτερίκευση της παραλλαγής Gln333 RV σε σύγκριση με την παραλλαγή άγριου τύπου. Η μετάλλαξη Asn194→Lys194 στον ιό RV G, η οποία εξηγεί την επανεμφάνιση του παθογόνου φαινοτύπου, συσχετίστηκε με σημαντική μείωση του χρόνου εσωτερίκευσης.[ 36 ] Επιπλέον, πειράματα με χιμαιρικούς RV έδειξαν ότι ο χρόνος που απαιτείται για την εσωτερίκευση των ιικών σωματιδίων του RV αυξήθηκε σημαντικά και η παθογένεια μειώθηκε σημαντικά μετά την αντικατάσταση του γονιδίου G του εξαιρετικά παθογόνου στελέχους SB RV, το οποίο προήλθε από έναν κλώνο cDNA του στελέχους RV-18 που σχετίζεται με νυχτερίδες και προέρχεται από άργυρο,[ 37 ] με αυτό του εξαιρετικά εξασθενημένου στελέχους SN, το οποίο απομονώθηκε από έναν κλώνο cDNA του στελέχους εμβολίου SAD B19 RV.[ 38 ] Μαζί, αυτά τα δεδομένα υποστηρίζουν την άποψη ότι η κινητική της πρόσληψης του ιού, η οποία είναι συνάρτηση του RV G, είναι ένας σημαντικός καθοριστικός παράγοντας της παθογένειας του RV.
- Ιικά στοιχεία που εμπλέκονται στην εξάπλωση και μετάδοση ιών
Μια μοναδική ιδιότητα του ιού της λύσσας είναι η ικανότητά του να εξαπλώνεται από κύτταρο σε κύτταρο. Η παρατήρηση ότι η παραλλαγή Gln333 ERA χάνει την εξαρτώμενη από το pH δραστικότητα σύντηξης κυττάρου-κυττάρου in vitro [ 39 ] και εμφανίζει σημαντικά μειωμένη ικανότητα εξάπλωσης από κύτταρο σε κύτταρο [ 40 ] υποδηλώνει ότι ο RV G παίζει επίσης βασικό ρόλο στην εξάπλωση από κύτταρο σε κύτταρο και, ως εκ τούτου, στη μετάδοση του ιού, πιθανώς μέσω της συντηξιογόνου δράσης του. Αυτή η πιθανότητα υποστηρίζεται περαιτέρω από το εύρημα ότι ο ρυθμός εξάπλωσης του παθογόνου SPBNGAK που αναστρέφει τον RV είναι σχεδόν διπλάσιος από αυτόν που προσδιορίστηκε για την μη παθογόνο παραλλαγή SPBNGA. Είναι ενδιαφέρον ότι η μετάλλαξη Asn 194 → Lys 194 στο G SPBNGAK προκάλεσε μετατόπιση του ορίου pH για τη σύντηξη μεμβράνης σε υψηλότερο pH, υποστηρίζοντας την υπόθεση ότι ένα υψηλότερο όριο pH για τη σύντηξη μεμβράνης σχετίζεται με αυξημένη εξάπλωση του ιού. [ 41 ]
Μελέτες διανευρωνικών δεικτών λοίμωξης από ιό RV σε αρουραίους [ 42 ] και πιθήκους rhesus [ 43 ] έχουν δείξει ότι ο ιός της λύσσας μεταναστεύει αποκλειστικά προς τα πίσω στους άξονες. Αν και αρκετές πρωτεΐνες του RV εμπλέκονται σε μηχανισμούς νευρωνικής μεταφοράς, η πρωτεΐνη G του RV φαίνεται να παίζει κυρίαρχο ρόλο στη διανευρωνική εξάπλωση της λοίμωξης από ιό RV. Για παράδειγμα, ενώ η περιφερική λοίμωξη με τον ιό της ιππικής λοιμώδους αναιμίας (EIAV) ψευδοτυποποιημένο με RV G οδηγεί σε ιική μεταφορά στον νωτιαίο μυελό, ο ίδιος EIAV ψευδοτυποποιημένος με τον ιό της κυστιδιακής στοματίτιδας G δεν κατάφερε να εισέλθει στο νευρικό σύστημα. [ 44 ] Επιπλέον, η ιική εξάπλωση του μεταλλαγμένου ERA G Arg 333 → Gln 333 στο ΚΝΣ βρέθηκε να είναι σημαντικά μειωμένη σε σύγκριση με το μεταλλαγμένο άγριου τύπου, υποδηλώνοντας περαιτέρω μια λειτουργία του άθικτου G του RV στην διασυναπτική εξάπλωση. Ωστόσο, τα πιο πειστικά στοιχεία για έναν σημαντικό ρόλο του RV G στη διασυναπτική μεταφορά προέρχονται από ενδοκρανιακή μόλυνση ποντικών με έναν ανασυνδυασμένο ιό RV με ανεπάρκεια G, η οποία έδειξε ότι η μόλυνση παρέμεινε περιορισμένη στους νευρώνες στο σημείο του εμβολιασμού χωρίς καμία ένδειξη εξάπλωσης σε δευτερογενείς νευρώνες.[ 45 ] Ωστόσο, είναι πιθανό ότι εκτός από το RV G, το RV M παίζει επίσης ρόλο στην εξάπλωση του ιού και επομένως στη διασυναπτική μεταφορά. Από αυτή την άποψη, αποδείχθηκε ότι η εξάπλωση της χιμαιρικής παραλλαγής SN-BMBG RV, η οποία περιέχει τόσο M όσο και G από το εξαιρετικά παθογόνο SB, ήταν σημαντικά υψηλότερη από την εξάπλωση της χιμαιρικής παραλλαγής SN-BG ή SN-BM, οι οποίες περιέχουν το G και το M από το SB, αντίστοιχα, υποδηλώνοντας ότι η βέλτιστη αλληλεπίδραση του M με το G μπορεί να παίζει σημαντικό ρόλο στην εξάπλωση του ιού από κύτταρο σε κύτταρο. [ 46 ] Δεδομένου ότι ο ιός RV M υποστηρίζει την εκβλάστηση του ιού, [ 47 ] είναι πιθανό η πιο αποτελεσματική εξάπλωση της χιμαιρικής παραλλαγής RV SN-BMBG να οφείλεται στη βέλτιστη εκβλάστηση του ιού στη μετασυναπτική μεμβράνη.
Πρόσφατες μελέτες έχουν δείξει ότι η αλληλεπίδραση μεταξύ του RV P και της ελαφριάς αλυσίδας δυνεΐνης συνδέει το RV RV RV με το σύστημα μεταφοράς κυττάρων ξενιστή, διευκολύνοντας έτσι την ανάδρομη αξονική μεταφορά του ιού.[ 48 ],[ 49 ] Ωστόσο, η περιφερική μόλυνση ενήλικων ποντικών έδειξε ότι η διαγραφή του τομέα δέσμευσης LC8 του RV P δεν εμποδίζει την είσοδο του ιού στο ΚΝΣ, υποδηλώνοντας ότι η πρωτεΐνη RV δεν εμπλέκεται άμεσα στην ανάδρομη αξονική εξάπλωση του RV.[ 50 ]
- Ιικά στοιχεία που ελέγχουν την αναπαραγωγή του ιού
Σε αντίθεση με πολλούς άλλους ιούς, όπως ο ιός της γρίπης, η παθογένεια του ιού RV είναι αντιστρόφως ανάλογη με τον ρυθμό σύνθεσης ιικού RNA και παραγωγής μολυσματικών ιικών σωματιδίων. Η σύγκριση των επιπέδων ιικού mRNA και γονιδιωματικού RNA που παράγονται από διαφορετικούς χιμαιρικούς ιούς υποδηλώνει ότι η μεταγραφή και η αντιγραφή του ιικού RNA ρυθμίζονται από πολλαπλούς παράγοντες, συμπεριλαμβανομένου του RV M, ο οποίος έχει αναγνωριστεί ως ένας παράγοντας trans-acting που μεσολαβεί στη μετάβαση από τα αρχικά υψηλά επίπεδα σύνθεσης mRNA στη σύνθεση γονιδιωματικού RNA.[ 51 ] Επιπλέον, το M από όλους τους ραβδοϊούς είναι ικανό να διακόψει την έκφραση ιικών γονιδίων συνδεόμενο με το RNP, με αποτέλεσμα τον σχηματισμό μιας εξαιρετικά συμπυκνωμένης δομής που μοιάζει με ραχοκοκαλιά, η οποία δεν είναι σε θέση να υποστηρίξει τη σύνθεση RNA.
Για τον εντοπισμό άλλων ιικών στοιχείων που ελέγχουν την παθογένεια ρυθμίζοντας την αντιγραφή του ιού, οι 5' τελικές αλληλουχίες του εξαιρετικά παθογόνου στελέχους SB αντικαταστάθηκαν σταδιακά με αλληλουχίες από το εξαιρετικά εξασθενημένο στέλεχος εμβολίου SN, με αποτέλεσμα τους ανασυνδυασμένους ιούς SB2 (τελική αλληλουχία [TS] + L), SB3 (TS + L + ψευδογονίδιο [Ψ]), SB4 (TS + L + Ψ + G) και SB5 (TS + L + Ψ + G + M). Η ενδομυϊκή μόλυνση με τους γονικούς ιούς SB και SN και τους χιμαιρικούς RVs SB2, SB3, SB4 και SB5 προκάλεσε τα υψηλότερα ποσοστά θνησιμότητας σε ποντίκια μολυσμένα με SB και καθόλου νοσηρότητα ή θνησιμότητα σε ποντίκια μολυσμένα με SN. Η αντικατάσταση των TS, L και SB με τα αντίστοιχα στοιχεία από το SN είχε ως αποτέλεσμα μια μέτρια μείωση της νοσηρότητας και της θνησιμότητας, και μια πρόσθετη ανταλλαγή G ή G συν M μείωσε σημαντικά ή εξάλειψε πλήρως την ιική παθογένεια.
Ο φαινοτυπικός χαρακτηρισμός αυτών των άγριου τύπου και χιμαιρικών RV σε ιστοκαλλιέργεια αποκάλυψε ότι η παθογένεια ενός δεδομένου RV συσχετίζεται αντιστρόφως ανάλογα με την ικανότητά του να αναπαράγεται σε νευρωνικά κύτταρα. Αν και το SB αναπαρήχθη σε επίπεδα σχεδόν 1000 φορές χαμηλότερα από το SN, και η αντικατάσταση των TS, L και στο SB από επίπεδα SN είχε μικρή επίδραση στην κινητική της ιικής ανάπτυξης, η πρόσθετη αντικατάσταση του G ή του G συν M του SB από τα αντίστοιχα γονίδια SN είχε ως αποτέλεσμα μια αύξηση 1-log στην παραγωγή ιού, υποδηλώνοντας ότι η κινητική της αντιγραφής του ιικού RNA, καθώς και η παραγωγή ιικών σωματιδίων ελέγχονται σε μεγάλο βαθμό από την πρωτεΐνη G του RV. Αυτό το συμπέρασμα υποστηρίζεται από δεδομένα που ελήφθησαν με παραλλαγές G του RV που διαφέρουν κατά ένα αμινοξύ στις πρωτεΐνες G τους. Η παθογόνος παραλλαγή του ιού της λύσσας SPBNGAK 194 παρήγαγε έναν τίτλο ιού σε κύτταρα NA που ήταν 1 λογαριθμικό χαμηλότερος από αυτόν που παρήγαγε η μη παθογόνος παραλλαγή SPBNGAN 194, και η ανάλυση PCR σε πραγματικό χρόνο έδειξε ότι οι ρυθμοί μεταγραφής και αντιγραφής του ιικού RNA σε κύτταρα NA μολυσμένα με SPBNGAK ήταν 5 και 10 φορές υψηλότεροι από ό,τι σε κύτταρα NA μολυσμένα με SPBNGAK.[ 52 ] Περαιτέρω ενδείξεις για μια αντίστροφη συσχέτιση μεταξύ της παθογένειας και του ρυθμού σύνθεσης ιικού RNA και παραγωγής ιικών σωματιδίων παρείχαν ποντίκια μολυσμένα με χιμαιρικούς ανασυνδυασμένους ιούς στους οποίους τα γονίδια G και M του εξασθενημένου στελέχους SN αντικαταστάθηκαν από εκείνα του εξαιρετικά παθογόνου στελέχους SB. Αυτά τα πειράματα αποκάλυψαν μια σημαντική αύξηση στην παθογένεια του γονικού στελέχους SN που έφερε το RV G σε σχέση με το παθογόνο στέλεχος SB. Η παθογένεια αυξήθηκε περαιτέρω όταν τόσο το G όσο και το M από το SB εισήχθησαν στο SN.
Η υποκατάσταση του G ή του M ή και των δύο στο SN με τα αντίστοιχα γονίδια από το SB συσχετίστηκε με σημαντική μείωση στον ρυθμό παραγωγής ιικών σωματιδίων καθώς και στον ρυθμό σύνθεσης ιικού RNA. Αυτά τα δεδομένα δείχνουν ότι τόσο το G όσο και το M παίζουν σημαντικό ρόλο στην παθογένεση του RV ρυθμίζοντας την αντιγραφή του ιού. Το εύρημα ότι η υποκατάσταση του G ή του G συν M στο SN με G ή G συν M του SB οδηγεί σε μέτρια έως ισχυρή μείωση στην μεταγραφή και την αντιγραφή του ιικού RNA, αντίστοιχα, ενώ η υποκατάσταση μόνο του M στο SN με M του SB οδηγεί σε ισχυρή αύξηση στην μεταγραφή και την αντιγραφή του ιικού RNA, δείχνει ότι το RV G έχει επίσης σημαντική ρυθμιστική λειτουργία στην μεταγραφή/αντιγραφή του ιικού RNA είτε μόνο του είτε μέσω αλληλεπίδρασης με την πρωτεΐνη Μ. Ο μηχανισμός με τον οποίο το γονίδιο RV G ελέγχει τη σύνθεση του ιικού RNA είναι άγνωστος. Ορισμένες αλληλουχίες νουκλεοτιδίων εντός των γονιδίων RV G, όπως αυτές που περιλαμβάνουν τα κωδικόνια για Arg333 και Lys 194, έχουν αναγνωριστεί ως στόχοι για τα κυτταρικά miRNAs. Έχει αποδειχθεί ότι η αναγνώριση στόχων από τα κυτταρικά miRNAs μπορεί να οδηγήσει σε θετική ή αρνητική ρύθμιση της ιικής αντιγραφής. [ 53 ] Οι υποκαταστάσεις Arg 333 → Glu 333 ή Lys 194 → Ser 194 εντός της αλληλουχίας γονιδίου G του ιού RV έχουν ως αποτέλεσμα την κατάργηση των αλληλουχιών-στόχων miRNA, η οποία με τη σειρά της σχετίζεται με σημαντική αύξηση του ρυθμού σύνθεσης ιικού RNA [Faber M, Πανεπιστήμιο Thomas Jefferson, PA, ΗΠΑ, Αδημοσίευτα Δεδομένα], υποδηλώνοντας ότι τα miRNA των κυττάρων του ξενιστή παίζουν επίσης σημαντικό ρόλο στη ρύθμιση της αντιγραφής του ιού RV, όπως έχει αποδειχθεί για άλλους ιούς RNA, συμπεριλαμβανομένου του ιού της κυστιδιακής στοματίτιδας και του HCV. [ 54 ], [ 55 ]
Η ρύθμιση της αντιγραφής του ιού φαίνεται να είναι ένας από τους σημαντικούς μηχανισμούς που εμπλέκονται στην παθογένεση του ιού RV. Για να αποφευχθεί η ανοσολογική απόκριση και να διατηρηθεί η ακεραιότητα του νευρωνικού δικτύου, τα παθογόνα στελέχη του ιού RV, αλλά όχι τα εξασθενημένα στελέχη, μπορούν να ρυθμίσουν τον ρυθμό ανάπτυξής τους. Ένας χαμηλότερος ρυθμός αντιγραφής πιθανότατα ωφελεί τα παθογόνα στελέχη του ιού RV διατηρώντας τη νευρωνική δομή που χρησιμοποιούν αυτοί οι ιοί για να φτάσουν στο ΚΝΣ. Μια άλλη εξήγηση για τον χαμηλότερο ρυθμό αντιγραφής του παθογόνου ιού RV είναι ότι, προκειμένου να αποφύγει την έγκαιρη ανίχνευση από το ανοσοποιητικό σύστημα του ξενιστή, ο ιός διατηρεί ελάχιστα επίπεδα έκφρασης των αντιγόνων του.
Σχέση μεταξύ έκφρασης G του ιού RV, απόπτωσης και παθογένειας
Είναι γνωστό ότι τα στελέχη του ιού της λύσσας του δρόμου που είναι σημαντικά πιο παθογόνα από τα στελέχη που έχουν προσαρμοστεί σε καλλιέργεια ιστών εκφράζουν πολύ περιορισμένα επίπεδα G και δεν προκαλούν απόπτωση μέχρι αργά στον κύκλο της μόλυνσης, γεγονός που υποδηλώνει ότι η παθογένεια ενός συγκεκριμένου στελέχους ιού συσχετίζεται αντιστρόφως ανάλογα με την έκφραση G του RV και την ικανότητα πρόκλησης απόπτωσης.[ 56 ] Άμεσες ενδείξεις για συσχέτιση μεταξύ του επιπέδου έκφρασης G και της έκτασης της απόπτωσης ελήφθησαν με τον ανασυνδυασμένο RV SPBNGA-GA, ο οποίος έφερε δύο πανομοιότυπα γονίδια G και υπερέκφρασε RV G.[ 57 ] Μορφολογικές μελέτες νευρωνικών καλλιεργειών που μολύνθηκαν με αυτόν τον ανασυνδυασμένο RV έδειξαν ότι ο κυτταρικός θάνατος αυξήθηκε σημαντικά παράλληλα με την υπερέκφραση G του RV και ότι η απόπτωση είναι ο κύριος μηχανισμός που εμπλέκεται στον θάνατο που προκαλείται από RV G. Συγκεκριμένα, η μείωση της χρώσης F-ακτίνης μετά από μόλυνση με SPBNGA-GA είναι συνεπής με τον αποπολυμερισμό των νηματίων ακτίνης που προκαλείται από απόπτωση. Επιπλέον, ο αριθμός των TUNEL-θετικών πυρήνων σε νευρώνες μολυσμένους με SPBNGA-GA αυξήθηκε σημαντικά σε σύγκριση με αυτόν στους μη μολυσμένους και στους νευρώνες μολυσμένους με SPBNGA. Ωστόσο, ο μηχανισμός με τον οποίο το γονίδιο G του ιού RV μεσολαβεί στη διαδικασία αποπτωτικής σηματοδότησης παραμένει σε μεγάλο βαθμό άγνωστος. Έχει προταθεί ότι η έκφραση G του ιού RV πάνω από ένα ορισμένο όριο διαταράσσει σοβαρά την κυτταρική μεμβράνη. Είναι πολύ πιθανό τα αποπτωτικά κύτταρα να μην αποβάλλονται γρήγορα στο ΚΝΣ και επομένως να υφίστανται δευτερογενή νέκρωση. [ 58 ] Από την άλλη πλευρά, η λοίμωξη από ιό RV και ιδιαίτερα η υπερέκφραση της πρωτεΐνης G του ιού RV μπορεί να οδηγήσει σε πυρόπτωση, μια οδό κυτταρικού θανάτου παρόμοια με την απόπτωση που, σε αντίθεση με την απόπτωση, περιλαμβάνει την ενεργοποίηση της κασπάσης 1 και έτσι οδηγεί σε νέκρωση. [ 59 ] Ο βαθμός νέκρωσης ή πυρόπτωσης που προκαλείται από τη λοίμωξη από ιό RV πιθανότατα παίζει κρίσιμο ρόλο στην πρόκληση αντιιικής ανοσίας. Ενώ τα αποπτωτικά κύτταρα διατηρούν την ακεραιότητα της μεμβράνης τους και δεν διεγείρουν την έμφυτη ανοσολογική απόκριση, τα νεκρωτικά κύτταρα γίνονται διαπερατά και εκκρίνουν ενδογενή ανοσοενισχυτικά που μπορούν να προκαλέσουν μια ισχυρή έμφυτη ανοσολογική απόκριση. [ 60 ]
Δεδομένου ότι το επίπεδο απόπτωσης/νέκρωσης συσχετίζεται με την ανοσογονικότητα του ιού RV, έχει προταθεί ότι η ανοσοδιεγερτική δράση των αποπτωτικών/νεκρωτικών κυττάρων πιθανότατα συμβάλλει στη δημιουργία προστατευτικής ανοσολογικής απόκρισης. Επομένως, η ρύθμιση της έκφρασης G του ιού RV είναι πιθανότατα ένας σημαντικός παράγοντας στην παθογένεση της λύσσας, καθώς παρέχει ένα μέσο για την επιβίωση και τη διασπορά παθογόνων παραλλαγών του ιού RV στο νευρικό σύστημα χωρίς να προκαλεί εμφανή νευρωνική βλάβη και να προκαλεί προστατευτική ανοσοαπόκριση που θα απέτρεπε τη μόλυνση.
Η έκφραση της G του ιού RV μπορεί να ρυθμίζεται στο επίπεδο της σύνθεσης RNA, στο μετα-μεταφραστικό επίπεδο ή και στα δύο. Τα επίπεδα της G του ιού RV που εκφράζονται από διαφορετικές χιμαιρικές παραλλαγές του ιού RV έχουν αποδειχθεί ότι αντικατοπτρίζονται από τον ρυθμό σύνθεσης ιικού RNA, γεγονός που υποδηλώνει ότι η διαφορική ρύθμιση της έκφρασης της G του ιού RV από αυτές τις παραλλαγές προκύπτει από διακυμάνσεις στον ρυθμό μεταγραφής του ιικού mRNA. Όπως και με τους ρυθμούς μεταγραφής του ιικού RNA, η ποσότητα της G του ιού RV που εκφράζεται από αυτές τις παραλλαγές συσχετίζεται αντιστρόφως με την ιική παθογένεια. Από την άλλη πλευρά, η μόλυνση πρωτογενών νευρωνικών καλλιεργειών με την λιγότερο παθογόνο παραλλαγή RV CVS-B2c είχε ως αποτέλεσμα τετραπλάσια υψηλότερα επίπεδα πρωτεΐνης G από τη μόλυνση με την εξαιρετικά παθογόνο παραλλαγή CVS-N2c, παρά τη σύνθεση συγκρίσιμων επιπέδων mRNA G και στις δύο μολύνσεις. Τα πειράματα παλμικής παρακολούθησης έδειξαν ότι τα υψηλότερα επίπεδα πρωτεΐνης G σε νευρώνες που έχουν μολυνθεί με CVS-B2c ήταν σε μεγάλο βαθμό αποτέλεσμα χαμηλότερου ρυθμού αποικοδόμησης της πρωτεΐνης G CVS-B2c σε σύγκριση με την πρωτεΐνη G CVS-N2c. Ωστόσο, ο μηχανισμός που οδηγεί στην ταχύτερη πρωτεολυτική αποικοδόμηση της πρωτεΐνης G CVS-N2c παραμένει προς διευκρίνιση.
Συμπτώματα λύσσα
Η περίοδος επώασης για τη λύσσα είναι κατά μέσο όρο 30-90 ημέρες. Σε περίπτωση μαζικής μόλυνσης μέσω μεγάλων τραυμάτων στο κεφάλι και το πρόσωπο, μπορεί να μειωθεί σε 12 ημέρες. Σε σπάνιες περιπτώσεις, η περίοδος επώασης μπορεί να διαρκέσει 1 έτος ή περισσότερο.
Υπάρχει μια αυστηρά διαδοχική αλλαγή τριών περιόδων της νόσου: πρόδρομη, διέγερση, παράλυση.
Η προδρομική περίοδος ξεκινά με την εμφάνιση πόνου ή τραβήγματος στο σημείο του δαγκώματος, καθώς και πόνου κατά μήκος των νεύρων. Στην περιοχή της ουλής, μπορεί να υπάρχει αίσθημα καύσου, κνησμός, μερικές φορές ερυθρότητα και πρήξιμο. Ο ασθενής εμφανίζει γενική αδιαθεσία, πονοκέφαλο, ναυτία. Παρατηρούνται έμετος, αύξηση της θερμοκρασίας του σώματος στους 37,5-38 ° C και συμπτώματα προοδευτικής ψυχικής διαταραχής: αυξημένη αντανακλαστική διέγερση, ανεξήγητο αίσθημα άγχους, φόβου, μελαγχολίας. Συχνά ο ασθενής είναι καταθλιπτικός, ανασταλτικός, αποσυρμένος, αρνείται να φάει, κοιμάται άσχημα, παραπονιέται για ζοφερές σκέψεις, τρομακτικά όνειρα. Η προδρομική περίοδος διαρκεί 2-3 ημέρες, μερικές φορές επεκτείνεται σε 7 ημέρες. Στο τέλος αυτής της περιόδου, μπορεί να υπάρξουν κρίσεις άγχους με βραχυπρόθεσμες δυσκολίες στην αναπνοή, αίσθημα σφιξίματος στο στήθος, συνοδευόμενες από ταχυκαρδία και αυξημένο αναπνευστικό ρυθμό.
Η περίοδος διέγερσης χαρακτηρίζεται από την εμφάνιση υδροφοβίας: όταν προσπαθεί να πιει, και στη συνέχεια στη θέα του νερού ή μιας υπενθύμισης αυτού, ο ασθενής βιώνει έναν σπασμωδικό σπασμό του φάρυγγα και του λάρυγγα, κατά τον οποίο πετάει την κούπα με νερό με μια κραυγή, πετάει μπροστά τρεμάμενα χέρια, πετάει πίσω το κεφάλι και το σώμα του. Ο λαιμός είναι τεντωμένος, μια επώδυνη γκριμάτσα παραμορφώνει το πρόσωπο, το οποίο γίνεται μπλε λόγω σπασμού των αναπνευστικών μυών. Τα μάτια διογκώνονται, εκφράζουν φόβο, ζητούν βοήθεια, οι κόρες είναι διασταλμένες, η εισπνοή είναι δύσκολη. Στο αποκορύφωμα της κρίσης, είναι πιθανή η καρδιακή και αναπνευστική ανακοπή. Η κρίση διαρκεί αρκετά δευτερόλεπτα, μετά την οποία η κατάσταση του ασθενούς φαίνεται να βελτιώνεται. Στη συνέχεια, κρίσεις σπασμών των μυών του λάρυγγα και του φάρυγγα μπορούν να εμφανιστούν ακόμη και από την κίνηση του αέρα (αεροφοβία), το έντονο φως (φωτοφοβία) ή μια δυνατή λέξη (ακουστικοφοβία). Οι κρίσεις συνοδεύονται από ψυχοκινητική διέγερση, κατά την οποία ο ασθενής συμπεριφέρεται σαν "τρελός". Η συνείδηση είναι θολωμένη κατά τη διάρκεια της κρίσης, αλλά καθαρίζει στην ενδιάμεση περίοδο. Κατά την περίοδο της διέγερσης, λόγω του αυξημένου τόνου του συμπαθητικού νευρικού συστήματος, οι ασθενείς εμφανίζουν απότομη αύξηση της σιελόρροιας (σιελόρροια) με αδυναμία κατάποσης σάλιου λόγω σπασμού των φαρυγγικών μυών. Ο ασθενής ψεκάζει το σάλιο. Μερικοί ασθενείς μπορεί να εμφανίσουν σημάδια μηνιγγισμού και ακόμη και οπισθότονου, και οι σπασμοί είναι συχνοί. Σε αυτή την περίπτωση, το εγκεφαλονωτιαίο υγρό μπορεί να μην αλλάξει, αλλά σε ορισμένους ασθενείς, η συγκέντρωση πρωτεΐνης μπορεί να αυξηθεί και ο αριθμός των κυττάρων μπορεί να αυξηθεί λόγω των λεμφοκυττάρων.
Χωρίς επαρκή θεραπεία, τα σημάδια αφυδάτωσης αυξάνονται, τα χαρακτηριστικά του προσώπου γίνονται πιο έντονα και το σωματικό βάρος μειώνεται. Η θερμοκρασία του σώματος αυξάνεται σε υψηλές τιμές. Είναι πιθανοί σπασμοί. Η διάρκεια του σταδίου διέγερσης είναι περίπου 2-3 ημέρες, σπάνια 4-5 ημέρες. Μια μοιραία έκβαση συμβαίνει συνήθως κατά τη διάρκεια μιας από τις κρίσεις. Σπάνια, ο ασθενής επιβιώνει μέχρι το τρίτο στάδιο της νόσου.
Κατά την περίοδο της παράλυσης, ο ασθενής ηρεμεί. Οι κρίσεις υδροφοβίας σταματούν, ο ασθενής μπορεί να πιει και να καταπιεί τροφή, η συνείδηση είναι καθαρή. Ωστόσο, παρά την φαινομενική ευεξία, σύντομα εμφανίζονται λήθαργος, απάθεια, κατάθλιψη, παράλυση των άκρων, πυελικές διαταραχές, παράλυση των κρανιακών νεύρων. Η θερμοκρασία του σώματος αυξάνεται στους 42-43 °C, η αρτηριακή πίεση μειώνεται και μέχρι το τέλος της πρώτης ημέρας επέρχεται θάνατος από παράλυση των καρδιαγγειακών και αναπνευστικών κέντρων.
Στο περιφερικό αίμα παρατηρούνται ουδετεροφιλική λευκοκυττάρωση, αυξημένη αιμοσφαιρίνη, ερυθροκύτταρα και αιματοκρίτης.
Τι σε προβληματιζει?
Έντυπα
Κλινικά, διακρίνονται οι τυπικές και οι άτυπες μορφές. Οι άτυπες μορφές περιλαμβάνουν όλες τις περιπτώσεις χωρίς διέγερση και υδροφοβία. Οι άτυπες μορφές περιλαμβάνουν την προμηλική, την παρεγκεφαλιδική, την μηνιγγοεγκεφαλίτιδα κ.λπ.
Διαγνωστικά λύσσα
Η ανίχνευση αντιγόνου λύσσας, αντισωμάτων, ιικού RNA ή απομόνωσης ιού επιτρέπει τη διάγνωση της λύσσας. Επειδή οποιαδήποτε μεμονωμένη εξέταση μπορεί να είναι αρνητική σε έναν ασθενή με λύσσα, μερικές φορές είναι απαραίτητα σειριακά δείγματα ορού για ανίχνευση αντισωμάτων λύσσας, δείγματα σάλιου για καλλιέργεια ιού και βιοψία δέρματος για άμεση ανοσοφθοριστική εξέταση για ιικό αντιγόνο, ειδικά όταν υπάρχει έντονη υποψία λύσσας.
Μία από τις πιο γρήγορες μεθόδους για τη διάγνωση της προθανάτιας λύσσας στους ανθρώπους είναι η διεξαγωγή άμεσης δοκιμής ανοσοφθορισμού σε βιοψία δέρματος του αυχένα για την ανίχνευση αντιγόνου λύσσας. Η δοκιμή άμεσου ανοσοφθορισμού είναι η πιο ευαίσθητη και ειδική μέθοδος για την ανίχνευση αντιγόνου λύσσας στο δέρμα και σε άλλους φρέσκους ιστούς (π.χ., βιοψία εγκεφάλου), αν και τα αποτελέσματα μπορεί περιστασιακά να είναι αρνητικά στα αρχικά στάδια της νόσου. Εάν δεν υπάρχει διαθέσιμος φρέσκος ιστός, η ενζυμική πέψη των σταθερών ιστών μπορεί να αυξήσει την αντιδραστικότητα της δοκιμής ανοσοφθορισμού. Ωστόσο, η ευαισθησία μπορεί να είναι απαράδεκτα χαμηλή.
Η διάγνωση μπορεί επίσης να τεθεί εάν ο ιός απομονωθεί από το σάλιο μετά από εμβολιασμό κυττάρων νευροβλαστώματος ή εργαστηριακών τρωκτικών. Αυτό είναι συνήθως πιο αποτελεσματικό κατά τη διάρκεια των πρώτων 2-3 εβδομάδων της ασθένειας. Η ανίχνευση αντισωμάτων εξουδετέρωσης του ιού της λύσσας, που συνήθως πραγματοποιείται με τη δοκιμή ταχείας αναστολής εστίασης φθορισμού (RFFIT), στον ορό μη εμβολιασμένων ατόμων είναι επίσης διαγνωστική. Η παρουσία αντισωμάτων στο εγκεφαλονωτιαίο υγρό επιβεβαιώνει τη διάγνωση, αλλά μπορεί να εμφανιστούν 2-3 ημέρες αργότερα από τα αντισώματα ορού και επομένως μπορεί να είναι λιγότερο χρήσιμα στα πρώιμα στάδια της νόσου. Ενώ η ορολογική απόκριση μετά τον εμβολιασμό είναι γενικά αδιάκριτη από την ορολογική απόκριση που προκαλείται από την ασθένεια, ο εμβολιασμός συνήθως δεν παράγει αντισώματα στο εγκεφαλονωτιαίο υγρό.
Μόνο επτά περιπτώσεις «ανάρρωσης» από λύσσα τα τελευταία 25 χρόνια έχουν τεκμηριωθεί επαρκώς. Παρόλο που ο ιός της λύσσας δεν απομονώθηκε από κανέναν από τους ασθενείς, οι υψηλοί τίτλοι εξουδετερωτικών αντισωμάτων κατά της λύσσας σε δείγματα ορού και η παρουσία εξουδετερωτικών αντισωμάτων στο εγκεφαλονωτιαίο υγρό υποστήριξαν έντονα τη διάγνωση.
Τι χρειάζεται να εξετάσετε;
Ποιες δοκιμές χρειάζονται;
Διαφορική διάγνωση
Η διάγνωση της λύσσας στον άνθρωπο γίνεται συνήθως με βάση επιδημιολογικά και κλινικά δεδομένα και επιβεβαιώνεται στο εργαστήριο. Η διάγνωση είναι απλή εάν υπάρχει ιστορικό δαγκωμάτων ζώων και έχει εμφανιστεί το πλήρες φάσμα των συμπτωμάτων και σημείων. Διαφορετικά, είναι απαραίτητη μια προσεκτική αλλά ταχεία αξιολόγηση των επιδημιολογικών και κλινικών χαρακτηριστικών λιγότερο τυπικών περιπτώσεων πριν από την εκτέλεση ειδικών εργαστηριακών εξετάσεων. Κάθε ασθενής με νευρολογικά σημεία ή συμπτώματα ή ανεξήγητη εγκεφαλίτιδα θα πρέπει να ερωτάται σχετικά με την πιθανότητα έκθεσης σε ζώα σε περιοχές ενδημικές της λύσσας εντός ή εκτός της χώρας διαμονής. Η μη υποψία λύσσας σε αρκετούς πρόσφατους θανάτους ανθρώπων στις Ηνωμένες Πολιτείες μπορεί να οφείλεται σε έλλειψη προσεκτικού ιστορικού έκθεσης.
Στην έναρξη της νόσου, η λύσσα μπορεί να μιμηθεί πολλές μολυσματικές και μη μολυσματικές ασθένειες. Πολλές άλλες εγκεφαλίτιδες, όπως αυτές που προκαλούνται από ερπητοϊούς και αρμποϊούς, μοιάζουν με τη λύσσα. Άλλες μολυσματικές ασθένειες μπορούν επίσης να μιμηθούν τη λύσσα, όπως ο τέτανος, η εγκεφαλική ελονοσία, η ρικετσιόζη και ο τυφοειδής πυρετός. Παραλυτικές μολυσματικές ασθένειες που μπορούν να συγχέονται με τη λύσσα περιλαμβάνουν την πολιομυελίτιδα, την αλλαντίαση και την ερπητική εγκεφαλίτιδα τύπου Β από πίθηκο έρπητα.
Μη μολυσματικές ασθένειες που μπορεί να συγχέονται με τη λύσσα περιλαμβάνουν μια σειρά από νευρολογικά σύνδρομα, ιδιαίτερα την οξεία φλεγμονώδη πολυνευροπάθεια (σύνδρομο Guillain-Barré), καθώς και την αλλεργική μετεμβολιαστική εγκεφαλομυελίτιδα δευτερογενή σε εμβολιασμό του νευρικού ιστού κατά της λύσσας, δηλητηρίαση ή φαρμακευτική δηλητηρίαση, στέρηση αλκοόλ, οξεία πορφυρία και υστερία λύσσας. Το σύνδρομο Guillain-Barré μπορεί να εκληφθεί εσφαλμένα ως παραλυτική λύσσα και αντίστροφα.
Ποιος θα επικοινωνήσει;
Θεραπεία λύσσα
Δεν έχει αναπτυχθεί θεραπεία για τη λύσσα. Η χορήγηση μεγάλων δόσεων ειδικής αντιλυσσικής ανοσοσφαιρίνης και λευκοκυττάρων ιντερφερόνης είναι αναποτελεσματική. Χορηγείται συμπτωματική θεραπεία για την ανακούφιση του πόνου του ασθενούς. Για το σκοπό αυτό, ο ασθενής τοποθετείται σε ξεχωριστό θάλαμο ή θάλαμο, δημιουργείται ένα προστατευτικό καθεστώς που περιορίζει την επίδραση του εξωτερικού περιβάλλοντος (μειωμένος θόρυβος, έντονο φως, ροή αέρα). Για τη μείωση της διέγερσης του κεντρικού νευρικού συστήματος, συνταγογραφούνται υπνωτικά χάπια, αντισπασμωδικά και παυσίπονα. Η ισορροπία του νερού ομαλοποιείται.
Στο παραλυτικό στάδιο, συνταγογραφούνται φάρμακα που διεγείρουν τη δραστηριότητα του καρδιαγγειακού και του αναπνευστικού συστήματος. Συνιστάται η χρήση υπερβαρικής οξυγόνωσης, εγκεφαλικής υποθερμίας, ελεγχόμενης μηχανικής αναπνοής με πλήρη κουραροποίηση του ασθενούς. Ωστόσο, όλες οι μέθοδοι θεραπείας είναι πρακτικά αναποτελεσματικές. Στην καλύτερη περίπτωση, είναι δυνατό να παραταθεί η ζωή του ασθενούς για αρκετούς μήνες. Ένα δυσμενές αποτέλεσμα προκαθορίζεται από τη σοβαρότητα της βλάβης στο εγκεφαλικό στέλεχος με την καταστροφή ζωτικών κέντρων.
Πρόληψη
Η ανάπτυξη του πρώτου εμβολίου κατά της λύσσας από τον Παστέρ το 1885 εγκαινίασε μια εποχή πολύ πιο αποτελεσματικού ελέγχου της λύσσας. Σήμερα, παρά το ποσοστό θνησιμότητας σχεδόν 100% στους ανθρώπους από λύσσα, η ασθένεια μπορεί να προληφθεί πλήρως μέσω εμβολιασμού πριν και/ή μετά την έκθεση. Ενώ ο Παστέρ και οι συνάδελφοί του ξεκίνησαν τον εμβολιασμό ιδιωτικών σκύλων στο Παρίσι, ο πρώτος μαζικός εμβολιασμός σκύλων πραγματοποιήθηκε στις αρχές της δεκαετίας του 1920 στην Ιαπωνία, σηματοδοτώντας το πρώτο σημαντικό εθνικό πρόγραμμα ελέγχου της λύσσας. Ο εμβολιασμός άγριων ζώων από το στόμα, που αναπτύχθηκε για πρώτη φορά τη δεκαετία του 1970, έχει έκτοτε επανειλημμένα αποδειχθεί ότι ελέγχει αποτελεσματικά την ασθένεια σε σημαντικούς χερσαίους ξενιστές όπως αλεπούδες, ρακούν και κουνάβια.[ 68 ] Ο παρατεταμένος εμβολιασμός κατά της λύσσας των πληθυσμών ζώων των δεξαμενών με ποσοστά κάλυψης 70% ή υψηλότερα θα εξαλείψει τελικά τον ιό RABV από τα είδη των δεξαμενών και θα αποτρέψει την εξάπλωση του ιού σε τυχαίους ξενιστές. [ 69 ]
Φυλογενετικά δεδομένα δείχνουν ότι οι ιοί της λύσσας μόλυναν τις νυχτερίδες πολύ πριν μολύνουν τα χερσαία θηλαστικά και οι περισσότεροι ιοί της λύσσας, συμπεριλαμβανομένου του ιού RABV, εξακολουθούν να κυκλοφορούν σε διάφορα είδη νυχτερίδων παγκοσμίως.[ 70 ] Ωστόσο, αποτελεσματικές μέθοδοι για την πρόληψη της μετάδοσης του ιού RABV μεταξύ των νυχτερίδων παραμένουν ασαφείς, αποκλείοντας την πιθανότητα πλήρους εξάλειψης της λύσσας προς το παρόν. Ωστόσο, ακόμη και μετά την έκθεση στον ιό RABV μέσω του δαγκώματος ενός θηλαστικού που έχει μολυνθεί από λύσσα, η ασφαλής και αποτελεσματική προφύλαξη μετά την έκθεση (PEP, συμπεριλαμβανομένου του καθαρισμού τραυμάτων, της ανοσοσφαιρίνης λύσσας και του εμβολιασμού κατά της λύσσας) μπορεί να προστατεύσει τους ανθρώπους από τη μόλυνση από λύσσα, εάν η θεραπεία χορηγηθεί άμεσα και σύμφωνα με τις συστάσεις του Παγκόσμιου Οργανισμού Υγείας (ΠΟΥ).
Αυτές οι δύο μέθοδοι πρόληψης ανθρώπινων θανάτων —η μία που βασίζεται στον εμβολιασμό των εκτεθειμένων ατόμων και η άλλη που βασίζεται στον εμβολιασμό επαρκούς αριθμού σκύλων για να σπάσει ο κύκλος μετάδοσης στην πηγή— αποτελούν τα δομικά στοιχεία μιας προσέγγισης «μίας υγείας» για την πρόληψη και τον έλεγχο της λύσσας στους σκύλους. Αυτά τα δύο διαφορετικά μέσα πρόληψης ανθρώπινων θανάτων εξετάστηκαν ως ξεχωριστές εναλλακτικές λύσεις: η Στρατηγική Α, που βασίζεται στην παροχή προληπτικής δράσης (PEP) στους ανθρώπους, και η Στρατηγική Β, που βασίζεται στον εμβολιασμό σκύλων· ή ως στοιχεία μιας συνδυασμένης Στρατηγικής Α + Β σε μια ανάλυση του πιθανού κόστους των εναλλακτικών στρατηγικών.[ 71 ]
Χώρες όπως η Ταϊλάνδη έχουν σημειώσει τεράστια επιτυχία στην πρόληψη των ανθρώπινων θανάτων μέσω της χρήσης PEP, αλλά έχουν επίσης διαπιστώσει αυξανόμενη ζήτηση και συναφές κόστος που σχετίζεται με τη χρήση μόνο της PEP. [ 72 ] Για παράδειγμα, σε σύγκριση με την κατάσταση το 1991, τέσσερις φορές περισσότεροι άνθρωποι (πάνω από 400.000) χρειάζονταν PEP το 2003. Πρόσφατα στοιχεία δείχνουν ότι η Λαϊκή Δημοκρατία της Κίνας, η οποία εμβολιάζει 15 εκατομμύρια ανθρώπους ετησίως μετά από πιθανή έκθεση στη λύσσα, δαπανά περίπου 650 εκατομμύρια δολάρια ΗΠΑ ετησίως μόνο για PEP. [ 73 ]
Μια πολύ πιο βιώσιμη προσέγγιση είναι η πρόληψη της εξάπλωσης της λοίμωξης στην πηγή, στον ζωικό πληθυσμό, αυξάνοντας παράλληλα την πρόσβαση στην PEP για τους εκτεθειμένους ανθρώπους ασθενείς όταν χρειάζεται. Όπου υπάρχει πολιτική βούληση και επαρκής χρηματοδότηση για τον έλεγχο της λύσσας στους σκύλους, οι θάνατοι μπορούν και έχουν εξαλειφθεί. Η ευρεία χρήση του εμβολιασμού των σκύλων έχει οδηγήσει στην εξάλειψη της λύσσας στους σκύλους από αρκετές χώρες, συμπεριλαμβανομένης της Μαλαισίας το 1954, [ 74 ] της Ιαπωνίας το 1956, της Ταϊβάν το 1961, της Σιγκαπούρης και, ειδικότερα, σε όλη τη Δυτική Ευρώπη (ανασκόπηση στο Rupprecht et al, King et al, και Gongal και Wright). [ 75 ]
Использованная литература