Ιατρικός εμπειρογνώμονας του άρθρου

Νέες δημοσιεύσεις
Ο ιός της ανθρώπινης ανοσοανεπάρκειας (HIV)
Τελευταία επισκόπηση: 04.07.2025

Όλα τα περιεχόμενα του iLive ελέγχονται ιατρικά ή ελέγχονται για να διασφαλιστεί η όσο το δυνατόν ακριβέστερη ακρίβεια.
Έχουμε αυστηρές κατευθυντήριες γραμμές προμήθειας και συνδέουμε μόνο με αξιόπιστους δικτυακούς τόπους πολυμέσων, ακαδημαϊκά ερευνητικά ιδρύματα και, όπου είναι δυνατόν, ιατρικά επισκοπικά μελέτες. Σημειώστε ότι οι αριθμοί στις παρενθέσεις ([1], [2], κλπ.) Είναι σύνδεσμοι με τις οποίες μπορείτε να κάνετε κλικ σε αυτές τις μελέτες.
Εάν πιστεύετε ότι κάποιο από το περιεχόμενό μας είναι ανακριβές, παρωχημένο ή αμφισβητήσιμο, παρακαλώ επιλέξτε το και πατήστε Ctrl + Enter.
Το σύνδρομο επίκτητης ανοσοανεπάρκειας αναγνωρίστηκε ως ειδική ασθένεια το 1981 στις Ηνωμένες Πολιτείες, όταν αρκετοί νέοι ανέπτυξαν σοβαρές ασθένειες που προκλήθηκαν από μικροοργανισμούς που δεν ήταν παθογόνοι ή ασθενώς παθογόνοι για τους υγιείς ανθρώπους. Μια μελέτη της ανοσολογικής κατάστασης των ασθενών αποκάλυψε μια απότομη μείωση στον αριθμό των λεμφοκυττάρων γενικά και των Τ-βοηθητικών λεμφοκυττάρων ειδικότερα. Αυτή η πάθηση ονομάστηκε AIDS (Σύνδρομο Επίκτητης Ανοσοανεπάρκειας). Η μέθοδος μόλυνσης (σεξουαλική επαφή, μέσω του αίματος και των παρασκευασμάτων του) υποδήλωνε τη μολυσματική φύση της ασθένειας.
Ο αιτιολογικός παράγοντας του AIDS ανακαλύφθηκε το 1983 ανεξάρτητα από τον Γάλλο L. Montagnier, ο οποίος τον ονόμασε ιό που σχετίζεται με τη λεμφαδενοπάθεια LAV, επειδή τον βρήκε σε έναν ασθενή με λεμφαδενοπάθεια, και τον Αμερικανό R. Gallo, ο οποίος ονόμασε τον ιό HTLV-III (Human T-lymphotropic Virus III): είχε προηγουμένως ανακαλύψει τους λεμφοτρόπους ιούς I και II.
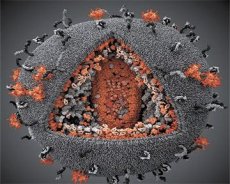
Μια σύγκριση των ιδιοτήτων των ιών LAV και HTLV-III έδειξε την ταυτότητά τους, έτσι για να αποφευχθεί η σύγχυση, ο ιός ονομάστηκε HIV (Ιός Ανθρώπινης Ανοσοανεπάρκειας ή HIV) το 1986. Ο HIV έχει σφαιρικό σχήμα, με διάμετρο 110 nm. Το περίβλημα του ιού έχει σχήμα πολυέδρου, που αποτελείται από 12 πεντάγωνα και 20 εξάγωνα. Στο κέντρο και στις γωνίες κάθε εξαγώνου υπάρχει ένα μόριο γλυκοζυλιωμένης πρωτεΐνης gpl20 (ο αριθμός 120 υποδεικνύει το μοριακό βάρος της πρωτεΐνης σε κιλοντάλτον). Συνολικά 72 μόρια gpl20 βρίσκονται στην επιφάνεια του ιού με τη μορφή ιδιόμορφων αιχμών, καθεμία από τις οποίες σχετίζεται με την ενδομεμβρανική πρωτεΐνη gp41. Αυτές οι πρωτεΐνες, μαζί με το διπλό λιπιδικό στρώμα, σχηματίζουν το υπερκαψίδιο (μεμβράνη) του ιού.
Οι πρωτεΐνες gpl20 και gp41 σχηματίζονται με την κοπή της πρόδρομης πρωτεΐνης Env από μια κυτταρική πρωτεάση. Η πρωτεΐνη gp41 σχηματίζει το "στέλεχος" της ακίδας συνδεόμενη με την κυτταροπλασματική της περιοχή στην πρωτεΐνη μήτρας p17MA που βρίσκεται ακριβώς κάτω από το περίβλημα. Τα μόρια p17 αλληλεπιδρούν κατά την ωρίμανση του ιικού σωματιδίου για να σχηματίσουν ένα εικοσάεδρο που βρίσκεται κάτω από το περίβλημα.
Στο κεντρικό τμήμα του ιικού σωματιδίου, η πρωτεΐνη p24 σχηματίζει ένα κωνικό καψίδιο. Το στενότερο τμήμα του καψιδίου συνδέεται με τη μεμβράνη του ιικού σωματιδίου με τη συμμετοχή της πρωτεΐνης rb. Μέσα στο καψίδιο, υπάρχουν δύο πανομοιότυπα μόρια ιικού γονιδιωματικού RNA. Συνδέονται μέσω των 5' άκρων τους με την πρωτεΐνη νουκλεοκαψιδίου p7NC. Αυτή η πρωτεΐνη είναι ενδιαφέρουσα επειδή έχει δύο υπολείμματα αμινοξέων (μοτίβα) πλούσια σε κυστεΐνη και ιστιδίνη και περιέχουν ένα άτομο Zn - ονομάζονται "δάκτυλα ψευδαργύρου" επειδή συλλαμβάνουν μόρια γονιδιωματικού RNA για να συμπεριληφθούν στα σχηματιζόμενα ιικά σωματίδια. Το καψίδιο περιέχει επίσης τρία ένζυμα. Η αναστροφάση (RT), ή σύμπλοκο pol, περιλαμβάνει την αντίστροφη μεταγραφάση, την RNase H και την DNA-εξαρτώμενη DNA πολυμεράση. Η αναστροφάση υπάρχει ως ετεροδιμερές p66/p51. Η πρωτεάση (PR) - p10, ξεκινά και εφαρμόζει τη διαδικασία ωρίμανσης του ιικού σωματιδίου. Η ιντεγκράση (IN) - p31, ή ενδονουκλεάση, διασφαλίζει την ενσωμάτωση του προϊικού DNA στο γονιδίωμα του κυττάρου ξενιστή. Το καψίδιο περιέχει επίσης ένα μόριο RNA εκκινητή (tRNA1"3).
Το γονιδίωμα RNA στο κύτταρο μετατρέπεται σε γονιδίωμα DNA (προϊός DNA) με τη βοήθεια της αντίστροφης μεταγραφάσης, που αποτελείται από 9283 ζεύγη νουκλεοτιδίων. Οριοθετείται αριστερά και δεξιά από τις λεγόμενες μακριές τερματικές επαναλήψεις ή LTR: S'-LTR στα αριστερά και 3'-LTR στα δεξιά. Οι LTR περιέχουν 638 ζεύγη νουκλεοτιδίων η καθεμία.
Το γονιδίωμα του HIV αποτελείται από 9 γονίδια, μερικά από τα οποία επικαλύπτονται στα άκρα (έχουν πολλά πλαίσια ανάγνωσης) και έχουν δομή εξονιντρονίου. Ελέγχουν τη σύνθεση 9 δομικών και 6 ρυθμιστικών πρωτεϊνών.
Η σημασία των LTR για το ιικό γονιδίωμα έγκειται στο ότι περιέχουν τα ακόλουθα ρυθμιστικά στοιχεία που ελέγχουν τη λειτουργία του:
- σήμα μεταγραφής (περιοχή υποκινητή)
- σήμα πρόσθεσης πολυ-Α;
- σήμα ανώτατου ορίου;
- σήμα ολοκλήρωσης;
- θετικό σήμα ρύθμισης (TAR για πρωτεΐνη TAT)·
- αρνητικό ρυθμιστικό στοιχείο (NRE για πρωτεΐνη NEF)·
- μια θέση για την προσκόλληση του εκκινητή RNA (tRNA™3) για τη σύνθεση DNA με αρνητική αλυσίδα στο 3' άκρο· ένα σήμα στο 5' άκρο του LTR που χρησιμεύει ως εκκινητής για τη σύνθεση DNA με θετική αλυσίδα.
Επιπλέον, η LTR περιέχει στοιχεία που εμπλέκονται στη ρύθμιση της συρραφής του mRNA, της συσκευασίας των μορίων vRNA στο καψίδιο (στοιχείο Psi). Τέλος, κατά τη διάρκεια της μεταγραφής του γονιδιώματος, σχηματίζονται δύο σήματα σε μακρά mRNA για την πρωτεΐνη REV, τα οποία αλλάζουν τη σύνθεση πρωτεϊνών: CAR - για ρυθμιστικές πρωτεΐνες και CRS - για δομικές πρωτεΐνες. Εάν η πρωτεΐνη REV συνδεθεί με την CAR, συντίθενται δομικές πρωτεΐνες. Εάν απουσιάζει, συντίθενται μόνο ρυθμιστικές πρωτεΐνες.
Τα ακόλουθα ρυθμιστικά γονίδια και οι πρωτεΐνες τους παίζουν ιδιαίτερα σημαντικό ρόλο στη ρύθμιση της λειτουργίας του γονιδιώματος του ιού:
- Πρωτεΐνη TAT, η οποία διεξάγει θετικό έλεγχο της ιικής αντιγραφής και δρα μέσω της ρυθμιστικής περιοχής TAR.
- Πρωτεΐνες NEV και VPU, οι οποίες ασκούν αρνητικό έλεγχο της αντιγραφής μέσω της περιοχής NRE.
- Πρωτεΐνη REV, η οποία διεξάγει θετικό-αρνητικό έλεγχο. Η πρωτεΐνη REV ελέγχει το έργο των γονιδίων gag, pol, env και διεξάγει αρνητική ρύθμιση του ματίσματος.
Έτσι, η αντιγραφή του HIV βρίσκεται υπό τριπλό έλεγχο - θετικό, αρνητικό και θετικά-αρνητικό.
Η πρωτεΐνη VIF καθορίζει τη μολυσματικότητα του νεοσυντιθέμενου ιού. Συνδέεται με την πρωτεΐνη καψιδίου p24 και υπάρχει στο ιικό σωματίδιο σε ποσότητα 60 μορίων. Η πρωτεΐνη NEF αντιπροσωπεύεται στο ιικό σωματίδιο από έναν μικρό αριθμό μορίων (5-10), πιθανώς συνδεδεμένων με το περίβλημα.
Η πρωτεΐνη VPR αναστέλλει τον κυτταρικό κύκλο στη φάση G2, συμμετέχει στη μεταφορά συμπλεγμάτων προενσωμάτωσης στον πυρήνα του κυττάρου, ενεργοποιεί ορισμένα ιικά και κυτταρικά γονίδια και αυξάνει την αποτελεσματικότητα της αντιγραφής του ιού σε μονοκύτταρα και μακροφάγα. Η θέση των πρωτεϊνών VPR, TAT, REV και VPU στο ιικό σωματίδιο δεν έχει προσδιοριστεί.
Εκτός από τις δικές της πρωτεΐνες, η μεμβράνη του ιού μπορεί να περιέχει ορισμένες πρωτεΐνες του κυττάρου ξενιστή. Οι πρωτεΐνες VPU και VPR συμμετέχουν στη ρύθμιση της αναπαραγωγής του ιού.
Αντιγονικές παραλλαγές του ιού της ανθρώπινης ανοσοανεπάρκειας (HIV)
Ο ιός της ανθρώπινης ανοσοανεπάρκειας (HIV) είναι πολύ μεταβλητός. Ακόμα και από το σώμα ενός ασθενούς, μπορούν να απομονωθούν στελέχη του ιού που διαφέρουν σημαντικά στις αντιγονικές ιδιότητες. Αυτή η μεταβλητότητα διευκολύνεται από την εντατική καταστροφή των κυττάρων CD4+ και μια ισχυρή αντισωματική απόκριση στη λοίμωξη από HIV. Μια νέα μορφή HIV, ο HIV-2, βιολογικά κοντά στον HIV-1 αλλά ανοσολογικά διαφορετική από αυτόν, έχει απομονωθεί από ασθενείς από τη Δυτική Αφρική. Η ομολογία της πρωτοταγούς δομής των γονιδιωμάτων αυτών των ιών είναι 42%. Το DNA του προϊού HIV-2 περιέχει 9671 bp και το LTR του - 854 bp. Ο HIV-2 στη συνέχεια απομονώθηκε σε άλλες περιοχές του κόσμου. Δεν υπάρχει διασταυρούμενη ανοσία μεταξύ HIV-1 και HIV-2. Δύο μεγάλες μορφές του HIV-1 είναι γνωστές: O (Outlier) και M (Major), ο τελευταίος διαιρείται σε 10 υποτύπους (AJ). Οκτώ υποτύποι (AH) κυκλοφορούν στη Ρωσία.
 [ 1 ], [ 2 ], [ 3 ], [ 4 ], [ 5 ], [ 6 ], [ 7 ], [ 8 ], [ 9 ], [ 10 ]
[ 1 ], [ 2 ], [ 3 ], [ 4 ], [ 5 ], [ 6 ], [ 7 ], [ 8 ], [ 9 ], [ 10 ]
Ο μηχανισμός αλληλεπίδρασης του HIV με το κύτταρο
Αφού διεισδύσει στο σώμα, ο ιός επιτίθεται πρώτα σε κύτταρα που περιέχουν τον ειδικό για αυτόν υποδοχέα CD4. Αυτός ο υποδοχέας υπάρχει σε μεγάλες ποσότητες στα Τ-βοηθητικά κύτταρα, σε μικρότερες ποσότητες στα μακροφάγα και τα μονοκύτταρα, και τα Τ-βοηθητικά κύτταρα είναι ιδιαίτερα ευαίσθητα στον ιό.
Ο ιός της ανθρώπινης ανοσοανεπάρκειας (HIV) αναγνωρίζει τους υποδοχείς CD4 χρησιμοποιώντας την πρωτεΐνη gpl20. Η διαδικασία αλληλεπίδρασης του HIV με το κύτταρο συμβαίνει σύμφωνα με το ακόλουθο σχήμα: προσρόφηση μέσω υποδοχέα -> επικαλυμμένο λάκκο -> επικαλυμμένο κυστίδιο -> λυσόσωμα. Σε αυτό, η μεμβράνη του ιού συντήκεται με τη μεμβράνη του λυσοσώματος και το πυρηνοκαψίδιο, απελευθερωμένο από το υπερκαψίδιο, εισέρχεται στο κυτταρόπλασμα. Στο δρόμο προς τον πυρήνα, καταστρέφεται και απελευθερώνεται το γονιδιωματικό RNA και τα σχετικά συστατικά του πυρήνα. Στη συνέχεια, η αντίστροφη μεταγραφάση συνθέτει την αρνητική αλυσίδα του DNA στο RNA του ιού, στη συνέχεια η RNase H καταστρέφει το RNA του ιού και η ιική DNA πολυμεράση συνθέτει την θετική αλυσίδα του DNA. 5'-LTR και 3'-LTR σχηματίζονται στα άκρα του DNA του προϊού. Ο DNA προϊός μπορεί να παραμείνει στον πυρήνα για κάποιο χρονικό διάστημα σε ανενεργή μορφή, αλλά αργά ή γρήγορα ενσωματώνεται στο χρωμόσωμα του κυττάρου-στόχου με τη βοήθεια της ιντεγκράσης του. Σε αυτό, ο προϊός παραμένει ανενεργός μέχρι το δεδομένο Τ-λεμφοκύτταρο να ενεργοποιηθεί από μικροβιακά αντιγόνα ή άλλα ανοσοεπαρκή κύτταρα. Η ενεργοποίηση της μεταγραφής του κυτταρικού DNA ρυθμίζεται από έναν ειδικό πυρηνικό παράγοντα (NF-kB). Είναι μια πρωτεΐνη που συνδέεται με το DNA και παράγεται σε μεγάλες ποσότητες κατά την ενεργοποίηση και τον πολλαπλασιασμό των Τ-λεμφοκυττάρων και των μονοκυττάρων. Αυτή η πρωτεΐνη συνδέεται με ορισμένες αλληλουχίες κυτταρικού DNA και παρόμοιες αλληλουχίες LTR του προϊού DNA και προκαλεί μεταγραφή τόσο του κυτταρικού DNA όσο και του προϊού DNA. Προκαλώντας μεταγραφή του προϊού DNA, πραγματοποιεί τη μετάβαση του ιού από μια ανενεργή κατάσταση σε μια ενεργή και, κατά συνέπεια, από μια επίμονη λοίμωξη σε μια παραγωγική. Ο προϊός μπορεί να παραμείνει σε ανενεργή κατάσταση για πολύ μεγάλο χρονικό διάστημα. Η ενεργοποίηση του ιού είναι μια κρίσιμη στιγμή στην αλληλεπίδρασή του με το κύτταρο.
Από τη στιγμή που ο ιός διεισδύει στο κύτταρο, ξεκινά η περίοδος της μόλυνσης από τον ιό HIV - η κατάσταση φορέα του ιού, η οποία μπορεί να διαρκέσει 10 χρόνια ή περισσότερο. και από τη στιγμή που ενεργοποιείται ο ιός, ξεκινά η ασθένεια - το AIDS. Με τη βοήθεια των ρυθμιστικών γονιδίων του και των προϊόντων τους, ο ιός αρχίζει να αναπαράγεται ενεργά. Η πρωτεΐνη TAT μπορεί να αυξήσει τον ρυθμό αναπαραγωγής του ιού κατά 1000 φορές. Η ιική μεταγραφή είναι πολύπλοκη. Περιλαμβάνει τον σχηματισμό τόσο πλήρους όσο και υπογονιδιωματικού mRNA, τη συρραφή mRNA και στη συνέχεια τη σύνθεση δομικών και ρυθμιστικών πρωτεϊνών.
Η σύνθεση των δομικών πρωτεϊνών γίνεται ως εξής. Αρχικά, συντίθεται η πρόδρομη πολυπρωτεΐνη Pr55Gag (μια πρωτεΐνη με μοριακό βάρος 55 kDa). Περιέχει 4 κύριες περιοχές: μήτρα (MA), καψίδιο (CA), νουκλεοκαψίδιο (NC) και την περιοχή rb, από την οποία, ως αποτέλεσμα της κοπής της Pr55Gag από την ιική πρωτεάση (αυτοαποκόπτεται από μια άλλη πρόδρομη πρωτεΐνη, την Gag-Pol), σχηματίζονται οι δομικές πρωτεΐνες p17, p24, p7 και rb, αντίστοιχα. Ο σχηματισμός της πολυπρωτεΐνης Pr55Gag είναι η κύρια προϋπόθεση για τον σχηματισμό ιικών σωματιδίων. Αυτή η πρωτεΐνη καθορίζει το πρόγραμμα μορφογένεσης του ιικού σωματιδίου. Περιλαμβάνει διαδοχικά τα στάδια μεταφοράς της πολυπρωτεΐνης Gag στην πλασματική μεμβράνη, την αλληλεπίδραση με αυτήν και τις αλληλεπιδράσεις πρωτεΐνης-πρωτεΐνης κατά τον σχηματισμό του ιικού σωματιδίου και την εκβλάστηση του. Η Pr55Gag συντίθεται σε ελεύθερα πολυριβοσώματα. Τα πρωτεϊνικά μόρια μεταφέρονται στη μεμβράνη, όπου αγκυρώνονται από τις υδρόφοβες περιοχές τους. Η περιοχή CA παίζει τον κύριο ρόλο στη δημιουργία της φυσικής διαμόρφωσης της πρωτεΐνης Gag. Η περιοχή NC εξασφαλίζει την ένταξη (με τη βοήθεια των "δακτύλων ψευδαργύρου") 2 μορίων γονιδιωματικού RNA στη σύνθεση του σχηματιζόμενου ιικού σωματιδίου. Το μόριο πολυπρωτεϊνης αρχικά διμερίζεται λόγω της αλληλεπίδρασης των περιοχών μήτρας. Στη συνέχεια, τα διμερή συνδυάζονται σε εξαμερή (6 μονάδων) σύμπλοκα ως αποτέλεσμα της αλληλεπίδρασης των περιοχών CA και NC. Τέλος, τα εξαμερή, ενώνοντας στις πλευρικές τους επιφάνειες, σχηματίζουν ανώριμα σφαιρικά ιοσωμάτια, μέσα στα οποία περιέχεται το γονιδιωματικό ιικό RNA, που συλλαμβάνεται από την περιοχή NC.
Μια άλλη πρόδρομη πρωτεΐνη, η Prl60Gag-Pol (μια πρωτεΐνη με μοριακό βάρος 160 kDa), συντίθεται ως αποτέλεσμα μιας μετατόπισης πλαισίου από το ριβόσωμα κατά τη μετάφραση του 3'-άκρου του γονιδίου gag στην περιοχή που βρίσκεται αμέσως πριν από την περιοχή που κωδικοποιεί την πρωτεΐνη rb. Αυτή η πολυπρωτεΐνη Gag-Pol περιέχει μια ατελή αλληλουχία πρωτεΐνης Gag (1-423 αμινοξέα) και αλληλουχίες Pol που περιλαμβάνουν τους τομείς PR, RT και IN. Τα μόρια πολυπρωτεΐνης Gag-Pol συντίθενται επίσης σε ελεύθερα πολυριβοσώματα και μεταφέρονται στην πλασματική μεμβράνη. Η πολυπρωτεΐνη Prl60Gagpol περιέχει όλες τις θέσεις διαμοριακής αλληλεπίδρασης και τις θέσεις δέσμευσης μεμβράνης που είναι εγγενείς στην πολυπρωτεΐνη Gag. Επομένως, τα μόρια πολυπρωτεΐνης Gag-Pol συντήκονται με τη μεμβράνη και, μαζί με τα μόρια Gag, περιλαμβάνονται στα σχηματιζόμενα ιικά σωματίδια, με αποτέλεσμα την εμφάνιση ενεργής πρωτεάσης και την έναρξη της διαδικασίας ωρίμανσης του ιικού σωματιδίου. Η πρωτεάση HIV-1 είναι ιδιαίτερα δραστική μόνο με τη μορφή διμερούς, επομένως, για την αυτοεκτομή της από την Prl60Gag-Pol, απαιτείται διμερισμός αυτών των μορίων. Η ωρίμανση του ιικού σωματιδίου συνίσταται στο γεγονός ότι η απελευθερούμενη ενεργή πρωτεάση κόβει την prl60Gag-Pol και την Gag55 στις θέσεις που αναγνωρίζει. Σχηματίζονται οι πρωτεΐνες p17, p24, p7, p6, η αναστροφάση, η ιντεγκράση και η σύνδεσή τους στη δομή του ιού.
Η πρωτεΐνη Env συντίθεται σε ριβοσώματα που σχετίζονται με τις μεμβράνες του ενδοπλασματικού δικτύου, στη συνέχεια γλυκοζυλιώνεται, κόβεται από μια κυτταρική πρωτεάση σε gp120 και gp41 και μεταφέρεται στην κυτταρική επιφάνεια. Σε αυτήν την περίπτωση, η gp41 διεισδύει στη μεμβράνη και συνδέεται με τις περιοχές μήτρας του μορίου πρωτεΐνης Gag που σχετίζονται με την εσωτερική επιφάνεια της μεμβράνης. Αυτή η σύνδεση διατηρείται στο ώριμο ιικό σωματίδιο.
Έτσι, η συναρμολόγηση των ιικών σωματιδίων συνίσταται στη συσσωμάτωση προδρόμων πρωτεϊνών και συναφών μορίων RNA στην πλασματική μεμβράνη του κυττάρου ξενιστή, στον σχηματισμό ανώριμων ιικών σωματιδίων και στην απελευθέρωσή τους με εκβλάστηση από την κυτταρική επιφάνεια. Κατά τη διάρκεια της εκβλάστησης, το ιικό σωματίδιο περιβάλλεται από μια κυτταρική μεμβράνη στην οποία ενσωματώνονται τα μόρια gp41 και gp120. Κατά τη διάρκεια της εκβλάστησης ή, ενδεχομένως, μετά την απελευθέρωση των ιικών σωματιδίων, συμβαίνει η ωρίμανσή τους, η οποία πραγματοποιείται με τη βοήθεια μιας ιικής πρωτεάσης και συνίσταται στην πρωτεολυτική κοπή των προδρόμων πρωτεϊνών Pr55Gag και Prl60Gag-Pol σε πρωτεΐνες του ώριμου ιού και στη σύνδεσή τους σε ορισμένα δομικά σύμπλοκα. Τον πρωταγωνιστικό ρόλο στις διαδικασίες της ιικής μορφογένεσης παίζει η πρόδρομη πολυπρωτεΐνη Pr55Gag, η οποία οργανώνει και συναρμολογεί το ανώριμο ιικό σωματίδιο. Η διαδικασία ωρίμανσής του ολοκληρώνεται από μια συγκεκριμένη ιική πρωτεάση.
Αιτίες ανοσοανεπάρκειας
Μία από τις κύριες αιτίες της ανοσοανεπάρκειας στη λοίμωξη από τον ιό HIV είναι ο μαζικός θάνατος των Τ-βοηθητικών λεμφοκυττάρων. Αυτό συμβαίνει ως αποτέλεσμα των ακόλουθων γεγονότων. Πρώτον, τα Τ-βοηθητικά λεμφοκύτταρα που έχουν μολυνθεί με τον ιό πεθαίνουν λόγω απόπτωσης. Πιστεύεται ότι σε ασθενείς με AIDS, η αντιγραφή του ιού, η απόπτωση και η μείωση του αριθμού των Τ-βοηθητικών λεμφοκυττάρων είναι αλληλένδετες. Δεύτερον, τα Τ-φονικά λεμφοκύτταρα αναγνωρίζουν και καταστρέφουν τα Τ-κύτταρα που έχουν μολυνθεί με τον ιό ή φέρουν προσροφημένα μόρια gpl20, καθώς και τα μολυσμένα με ιό και τα μη μολυσμένα με ιό Τ-βοηθητικά λεμφοκύτταρα, τα οποία σχηματίζουν συμπλάστες (συγκύτιο) που αποτελούνται από αρκετές δεκάδες κύτταρα (μερικά από αυτά πεθαίνουν ως αποτέλεσμα της αναπαραγωγής ιών σε αυτά). Ως αποτέλεσμα της καταστροφής ενός μεγάλου αριθμού Τ-βοηθητικών υποδοχέων, παρατηρείται μείωση της έκφρασης των μεμβρανικών υποδοχέων στα Β-λεμφοκύτταρα σε ιντερλευκίνη-2, η σύνθεση διαφόρων ιντερλευκινών (παράγοντες ανάπτυξης και διαφοροποίηση των Β-λεμφοκυττάρων - IL-4, IL-5, IL-6, κ.λπ.) διαταράσσεται, με αποτέλεσμα τη διαταραχή της λειτουργίας του συστήματος Τ-δολοφόνων. Η δραστηριότητα των συστημάτων συμπληρώματος και μακροφάγων καταστέλλεται. Τα μακροφάγα και τα μονοκύτταρα που έχουν μολυνθεί με τον ιό δεν πεθαίνουν για μεγάλο χρονικό διάστημα, αλλά δεν είναι σε θέση να απομακρύνουν τον ιό από το σώμα. Τέλος, λόγω της δομικής και αντιγονικής ομοιότητας της gpl20 με τους υποδοχείς ορισμένων επιθηλιακών κυττάρων του σώματος (συμπεριλαμβανομένων των υποδοχέων τροφοβλαστών που μεσολαβούν στη μετάδοση του HIV μέσω μοσχεύματος), συντίθενται αντισώματα αντι-υποδοχέα με ευρύ φάσμα δράσης. Τέτοια αντισώματα είναι ικανά να μπλοκάρουν διάφορους κυτταρικούς υποδοχείς και να περιπλέξουν την πορεία της νόσου με αυτοάνοσες διαταραχές. Η συνέπεια της λοίμωξης από HIV είναι η ήττα όλων των κύριων κρίκων του ανοσοποιητικού συστήματος. Τέτοιοι ασθενείς γίνονται ανυπεράσπιστοι έναντι μιας ευρείας ποικιλίας μικροοργανισμών. Αυτό οδηγεί στην ανάπτυξη ευκαιριακών λοιμώξεων και όγκων. Για τους ασθενείς με λοίμωξη από HIV, αυξάνεται ο κίνδυνος ανάπτυξης τουλάχιστον τριών τύπων καρκίνου: σάρκωμα Kaposi, καρκίνωμα (συμπεριλαμβανομένου του καρκίνου του δέρματος), λέμφωμα Β-κυττάρων, το οποίο εμφανίζεται λόγω κακοήθους μετασχηματισμού των Β-λεμφοκυττάρων. Ωστόσο, ο HIV δεν είναι μόνο λεμφοκυτταροτρόπος, αλλά και νευροτρόπος. Διεισδύει στα κύτταρα του κεντρικού νευρικού συστήματος (αστροκύτταρα) τόσο μέσω ενδοκυττάρωσης που προκαλείται από υποδοχείς όσο και μέσω φαγοκυττάρωσης λεμφοβλαστών που έχουν μολυνθεί από ιό από αστροκύτταρα. Όταν ο ιός αλληλεπιδρά με αστροκύτταρα, σχηματίζονται επίσης σύμπλαστοι, οι οποίοι διευκολύνουν την εξάπλωση του παθογόνου μέσω των μεσοκυττάριων καναλιών. Ο ιός μπορεί να παραμείνει σε μακροφάγα και μονοκύτταρα για μεγάλο χρονικό διάστημα, επομένως χρησιμεύουν ως δεξαμενή και διανομέας του στο σώμα, όντας σε θέση να διεισδύσουν σε όλους τους ιστούς. Τα μολυσμένα μακροφάγα παίζουν σημαντικό ρόλο στην εισαγωγή του HIV στο κεντρικό νευρικό σύστημα και στη βλάβη του. Στο 10% των ασθενών, τα πρωτοπαθή κλινικά σύνδρομα σχετίζονται με βλάβη στο κεντρικό νευρικό σύστημα και εκδηλώνονται ως άνοια. Έτσι, τα άτομα που έχουν μολυνθεί με HIV χαρακτηρίζονται από 3 ομάδες ασθενειών - ευκαιριακές λοιμώξεις,νεοπλασματικές ασθένειες και βλάβες στο κεντρικό νευρικό σύστημα.
Επιδημιολογία της λοίμωξης από HIV
Η πηγή της λοίμωξης από τον ιό HIV είναι μόνο ένα άτομο - ένα άρρωστο άτομο ή ένας φορέας του ιού. Ο ιός της ανθρώπινης ανοσοανεπάρκειας (HIV) περιέχεται στο αίμα, το σπέρμα, το τραχηλικό υγρό· στις θηλάζουσες μητέρες - στο μητρικό γάλα. Η μόλυνση γίνεται σεξουαλικά, μέσω του αίματος και των παρασκευασμάτων του, καθώς και από τη μητέρα στο παιδί πριν, κατά τη διάρκεια και μετά τον τοκετό. Δεν είναι γνωστές περιπτώσεις μόλυνσης από τον ιό μέσω τροφών, ποτών και τσιμπημάτων εντόμων.
Ο εθισμός στα ναρκωτικά συμβάλλει στην εξάπλωση του AIDS. Η συχνότητα εμφάνισης του HIV αυξάνεται κάθε χρόνο. Σύμφωνα με τον ΠΟΥ, από το 1980 έως το 2000, 58 εκατομμύρια άνθρωποι μολύνθηκαν από τον ιό HIV. Μόνο το 2000, 5,3 εκατομμύρια άνθρωποι μολύνθηκαν παγκοσμίως και 3 εκατομμύρια άνθρωποι πέθαναν από AIDS. Από την 1η Ιανουαρίου 2004, 264 χιλιάδες άτομα μολυσμένα με HIV είχαν καταγραφεί στη Ρωσία. Τα μισά από τα άτομα που έχουν μολυνθεί με HIV πεθαίνουν εντός 11-12 ετών από τη στιγμή της μόλυνσης. Στις αρχές του 2004, από κάθε 100 χιλιάδες πολίτες της Ρωσίας, περίπου 180 ζούσαν με διάγνωση «λοίμωξης HIV». Προβλέπεται ότι με αυτό το επίπεδο νοσηρότητας, ο συνολικός αριθμός των ατόμων που έχουν μολυνθεί από τον ιό HIV στη Ρωσία έως το 2012 θα είναι 2,5-3 εκατομμύρια άνθρωποι. Η πολυπλοκότητα της καταπολέμησης της λοίμωξης από τον ιό HIV εξαρτάται από διάφορους λόγους: πρώτον, δεν υπάρχουν αποτελεσματικές μέθοδοι θεραπείας και ειδικής πρόληψης. Δεύτερον, η περίοδος επώασης για τη λοίμωξη από τον ιό HIV μπορεί να υπερβεί τα 10 χρόνια. Η διάρκειά του εξαρτάται από τη στιγμή ενεργοποίησης του Τ-λεμφοκυττάρου και του DNA του προϊού που περιέχεται στο χρωμόσωμά του. Δεν είναι ακόμη σαφές εάν όλοι όσοι έχουν μολυνθεί με τον ιό είναι καταδικασμένοι σε AIDS ή εάν είναι δυνατή η μακροχρόνια μεταφορά του ιού χωρίς ασθένεια (κάτι που φαίνεται απίθανο). Τέλος, υπάρχουν αρκετοί ιοί ανθρώπινης ανοσοανεπάρκειας (HIV-1, HIV-2), οι αντιγονικές διαφορές μεταξύ των οποίων εμποδίζουν τον σχηματισμό διασταυρούμενης ανοσίας. Η ανακάλυψη του ιού της ανοσοανεπάρκειας των πιθήκων (SIV) ρίχνει φως στο ζήτημα της προέλευσης του HIV. Ο SIV είναι παρόμοιος με τον HIV στην οργάνωση του γονιδιώματός του, αλλά διαφέρει σημαντικά στην αλληλουχία νουκλεοτιδίων του. Ο HIV-2 καταλαμβάνει μια ενδιάμεση θέση μεταξύ του HIV-1 και του SIV στις ορολογικές του ιδιότητες και είναι πιο κοντά στον SIV στην αλληλουχία νουκλεοτιδίων του. Από αυτή την άποψη, ο VM Zhdanov πρότεινε ότι οι ιοί HIV-1, HIV-2 και SIV προήλθαν από έναν κοινό πρόγονο. Είναι πιθανό, σύμφωνα με τον R. Gallo, ότι ένας από τους SIV εισήλθε με κάποιο τρόπο στο ανθρώπινο σώμα, όπου υπέστη μια σειρά μεταλλάξεων, με αποτέλεσμα την εμφάνιση του HIV-1, του HIV-2 και των άλλων μορφών του.
Συμπτώματα της λοίμωξης από HIV
Ο ιός της ανθρώπινης ανοσοανεπάρκειας έχει ορισμένα χαρακτηριστικά που καθορίζουν σε μεγάλο βαθμό την παθογένεση της νόσου. Ο ιός έχει πολύ υψηλό ρυθμό αναπαραγωγής, ο οποίος καθορίζεται από τα ρυθμιστικά του στοιχεία (έως 5.000 ιικά σωματίδια συντίθενται σε 5 λεπτά στο ενεργό στάδιο). Λόγω της παρουσίας της πρωτεΐνης σύντηξης (gp41), ο ιός προκαλεί τον σχηματισμό εκτεταμένων συγκυτιακών δομών λόγω της σύντηξης μολυσμένων και μη μολυσμένων Τ-βοηθητικών, η οποία έχει ως αποτέλεσμα τον μαζικό θάνατό τους. Τα μόρια πρωτεΐνης gpl20 που σχηματίζονται σε μεγάλες ποσότητες κυκλοφορούν ελεύθερα στο αίμα και συνδέονται με τους υποδοχείς των μη μολυσμένων Τ-βοηθητικών, με αποτέλεσμα να αναγνωρίζονται και να καταστρέφονται από Τ-δολοφόνους. Ο ιός μπορεί να εξαπλωθεί μέσω μεσοκυττάριων καναλιών από κύτταρο σε κύτταρο, οπότε καθίσταται δυσπρόσιτος στα αντισώματα.
Κλινικά κριτήρια για τη λοίμωξη από HIV
Στους ενήλικες, η λοίμωξη από τον ιό HIV διαγιγνώσκεται όταν έχουν τουλάχιστον δύο σοβαρά συμπτώματα σε συνδυασμό με τουλάχιστον ένα ήπιο σύμπτωμα και απουσία άλλων γνωστών αιτιών ανοσοανεπάρκειας (καρκίνος, συγγενής ανοσοανεπάρκεια, σοβαρή ασιτία κ.λπ.). Τα σοβαρά συμπτώματα περιλαμβάνουν:
- απώλεια βάρους 10% ή περισσότερο·
- μια παρατεταμένη εμπύρετη κατάσταση, διαλείπουσα ή συνεχής.
- χρόνια διάρροια.
Ήπια συμπτώματα: επίμονος βήχας, γενικευμένη δερματίτιδα, υποτροπιάζων έρπης ζωστήρας, στοματική και φαρυγγική καντιντίαση, χρόνιος απλός έρπης, γενικευμένη λεμφαδενοπάθεια. Η διάγνωση του AIDS τίθεται μόνο με την παρουσία σαρκώματος Kaposi, κρυπτοκοκκικής μηνιγγίτιδας, πνευμονιοκυστικής πνευμονίας. Η κλινική εικόνα της νόσου επηρεάζεται από τη συνοδό ευκαιριακή λοίμωξη.
Μέθοδοι καλλιέργειας του ιού της ανθρώπινης ανοσοανεπάρκειας (HIV)
Οι HIV-1 και HIV-2 μπορούν να καλλιεργηθούν σε κύτταρα μόνο ενός κλώνου λεμφοκυττάρων TCV4 - H9, που λαμβάνεται από λευχαιμικά λεμφοκύτταρα TCV4. Για τους σκοπούς αυτούς μπορούν επίσης να χρησιμοποιηθούν μονοστρωματικές καλλιέργειες αστροκυττάρων, στα οποία ο HIV-1 αναπαράγεται καλά. Οι χιμπατζήδες είναι ευαίσθητοι στον HIV-1 μεταξύ των ζώων.
Η αντοχή του ιού στο εξωτερικό περιβάλλον είναι χαμηλή. Νεκρώνεται υπό την επίδραση του ηλιακού φωτός και της υπεριώδους ακτινοβολίας, καταστρέφεται στους 80 °C εντός 30 λεπτών, όταν υποβάλλεται σε επεξεργασία με συνήθως χρησιμοποιούμενα απολυμαντικά - εντός 20-30 λεπτών. Για την απολύμανση υλικού που περιέχει ιό, είναι απαραίτητο να χρησιμοποιηθούν μυκοβακτηριοκτόνα απολυμαντικά, καθώς είναι αποτελεσματικά έναντι μικροοργανισμών με την υψηλότερη αντοχή.
Εργαστηριακή διάγνωση της λοίμωξης από HIV
Η κύρια μέθοδος για τη διάγνωση της μεταφοράς του ιού και της λοίμωξης από HIV είναι η ενζυμική ανοσοδοκιμασία. Ωστόσο, λόγω του γεγονότος ότι η gpl20 έχει δομική και αντιγονική ομοιότητα με τους υποδοχείς ορισμένων ανθρώπινων κυττάρων, συμπεριλαμβανομένων των υποδοχέων που μεταφέρουν ανοσοσφαιρίνες μέσω των επιθηλιακών κυττάρων των βλεννογόνων, αντισώματα που σχετίζονται με αντισώματα κατά της gpl20 μπορεί να εμφανιστούν στο σώμα. Σε αυτήν την περίπτωση, μπορεί να υπάρχουν ψευδώς θετικά αποτελέσματα ELISA. Επομένως, όλοι οι θετικά αντιδρώντες οροί των ατόμων υποβάλλονται σε πρόσθετη ανάλυση χρησιμοποιώντας τη μέθοδο ανοσοαποτύπωσης ή Western blotting. Αυτή η μέθοδος βασίζεται στην ταυτοποίηση των υπό μελέτη αντισωμάτων μετά τον ηλεκτροφορητικό διαχωρισμό τους και την επακόλουθη δοκιμή χρησιμοποιώντας επισημασμένα αντισώματα κατά του είδους. Η ιολογική μέθοδος σπάνια χρησιμοποιείται λόγω της πολυπλοκότητας της καλλιέργειας του ιού. Ο κλώνος λεμφοκυττάρων H9 χρησιμοποιείται για την απόκτηση ιικών αντιγόνων - τα απαραίτητα συστατικά των διαγνωστικών συστημάτων δοκιμών. Η μέθοδος CPR επιτρέπει την ανίχνευση του ιού σε πρώιμο στάδιο της ιαιμίας.
Θεραπεία της λοίμωξης από HIV
Είναι απαραίτητο να βρεθούν ή να συντεθούν φάρμακα που καταστέλλουν αποτελεσματικά τη δράση της αντίστροφης μεταγραφάσης (ρεβερτάσης) ή της ιικής πρωτεάσης. Θα εμπόδιζαν τον σχηματισμό DNA προϊού και (ή) θα ανέστειλαν την ενδοκυτταρική αναπαραγωγή του ιού. Η σύγχρονη στρατηγική για τη θεραπεία ασθενών που έχουν μολυνθεί από τον ιό HIV βασίζεται στην αρχή της συνδυασμένης χρήσης φαρμάκων που αναστέλλουν την ιική πρωτεάση (ένα από τα φάρμακα) και την αντίστροφη δράση (2 διαφορετικά φάρμακα) - συνδυασμένη (τριπλή) θεραπεία. Στη Ρωσία, για τη θεραπεία ασθενών που έχουν μολυνθεί από τον ιό HIV, συνιστάται η συνδυασμένη χρήση 2 εγχώριων φαρμάκων: φωσφαζίδη και κριξιβάνη, τα οποία καταστέλλουν ειδικά την αναπαραγωγή του HIV σε πρώιμα και όψιμα στάδια αναπαραγωγής, ειδικά με μειωμένη δράση της αζιδοθυμιδίνης.
Το πρόβλημα της ειδικής προφύλαξης είναι η ανάγκη δημιουργίας ενός εμβολίου που θα εξασφάλιζε τον σχηματισμό αποτελεσματικής κυτταρικής ανοσίας βασισμένης σε κυτταροτοξικά λεμφοκύτταρα ειδικά για τον ιό χωρίς σημαντική παραγωγή αντισωμάτων. Αυτή η ανοσία παρέχεται από βοηθούς Thl. Είναι πιθανό τα αντισώματα, συμπεριλαμβανομένων των εξουδετερωτικών του ιού, όχι μόνο να είναι αναποτελεσματικά στην καταστολή της λοίμωξης HIV, αλλά σε υψηλό επίπεδο καταστέλλουν την κυτταρική ανοσία. Επομένως, ένα εμβόλιο κατά του HIV πρέπει να πληροί, πρώτα απ 'όλα, δύο κύριες απαιτήσεις: α) να είναι απολύτως ασφαλές και β) να διεγείρει τη δραστηριότητα των Τ-κυτταροτοξικών λεμφοκυττάρων. Μελετάται η αποτελεσματικότητα διαφόρων παραλλαγών εμβολίου που λαμβάνονται από νεκρούς (αδρανοποιημένους) ιούς και από μεμονωμένα αντιγόνα με υψηλές προστατευτικές ιδιότητες. Τέτοια αντιγόνα μπορούν είτε να απομονωθούν από τα ίδια τα ιοσωμάτια είτε να συντεθούν χημικά. Έχει προταθεί ένα εμβόλιο που δημιουργείται με βάση μεθόδους γενετικής μηχανικής. Πρόκειται για έναν ανασυνδυασμένο ιό vaccinia που φέρει γονίδια HIV υπεύθυνα για τη σύνθεση αντιγόνων με ισχυρές ανοσογόνες ιδιότητες. Το ζήτημα της αποτελεσματικότητας αυτών των εμβολίων απαιτεί σημαντικό χρόνο λόγω της μακράς περιόδου επώασης της λοίμωξης HIV και της υψηλής μεταβλητότητας του παθογόνου. Η δημιουργία ενός εξαιρετικά αποτελεσματικού εμβολίου κατά του HIV αποτελεί ένα επείγον θεμελιώδες πρόβλημα.