Ιατρικός εμπειρογνώμονας του άρθρου

Νέες δημοσιεύσεις
Vibrio της χολέρας
Τελευταία επισκόπηση: 04.07.2025

Όλα τα περιεχόμενα του iLive ελέγχονται ιατρικά ή ελέγχονται για να διασφαλιστεί η όσο το δυνατόν ακριβέστερη ακρίβεια.
Έχουμε αυστηρές κατευθυντήριες γραμμές προμήθειας και συνδέουμε μόνο με αξιόπιστους δικτυακούς τόπους πολυμέσων, ακαδημαϊκά ερευνητικά ιδρύματα και, όπου είναι δυνατόν, ιατρικά επισκοπικά μελέτες. Σημειώστε ότι οι αριθμοί στις παρενθέσεις ([1], [2], κλπ.) Είναι σύνδεσμοι με τις οποίες μπορείτε να κάνετε κλικ σε αυτές τις μελέτες.
Εάν πιστεύετε ότι κάποιο από το περιεχόμενό μας είναι ανακριβές, παρωχημένο ή αμφισβητήσιμο, παρακαλώ επιλέξτε το και πατήστε Ctrl + Enter.
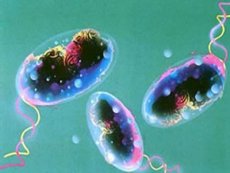
Σύμφωνα με τον ΠΟΥ, η χολέρα είναι μια μολυσματική ασθένεια που χαρακτηρίζεται από οξεία, σοβαρή, αφυδατωτική διάρροια με κόπρανα με τη μορφή νερού ρυζιού, η οποία είναι συνέπεια της μόλυνσης από Vibrio cholerae. Λόγω του γεγονότος ότι χαρακτηρίζεται από έντονη ικανότητα ευρείας εξάπλωσης σε επιδημίες, σοβαρή πορεία και υψηλό ποσοστό θνησιμότητας, η χολέρα θεωρείται ιδιαίτερα επικίνδυνη λοίμωξη.
Η ιστορική πατρίδα της χολέρας είναι η Ινδία, ή πιο συγκεκριμένα, το δέλτα των ποταμών Γάγγη και Βραχμαπούτρα (τώρα Ανατολική Ινδία και Μπαγκλαντές), όπου υπάρχει από αμνημονεύτων χρόνων (επιδημίες χολέρας σε αυτήν την περιοχή παρατηρήθηκαν ήδη από το 500 π.Χ.). Η μακρά ύπαρξη μιας ενδημικής εστίας χολέρας εδώ εξηγείται από πολλούς λόγους. Το δονίδιο χολέρας μπορεί όχι μόνο να επιβιώσει στο νερό για μεγάλο χρονικό διάστημα, αλλά και να αναπαραχθεί σε αυτό υπό ευνοϊκές συνθήκες - θερμοκρασίες άνω των 12 °C, παρουσία οργανικής ύλης. Όλες αυτές οι συνθήκες είναι εμφανείς στην Ινδία: τροπικό κλίμα (μέση ετήσια θερμοκρασία από 25 έως 29 °C), άφθονες βροχοπτώσεις και βάλτος, υψηλή πυκνότητα πληθυσμού, ειδικά στο δέλτα του ποταμού Γάγγη, μεγάλη ποσότητα οργανικής ύλης στο νερό, συνεχής ρύπανση των υδάτων όλο το χρόνο με λύματα και περιττώματα, χαμηλό υλικό βιοτικό επίπεδο και μοναδικές θρησκευτικές και λατρευτικές τελετουργίες του πληθυσμού.
Στην ιστορία των επιδημιών χολέρας, μπορούν να διακριθούν τέσσερις περίοδοι.
Περίοδος Ι - μέχρι το 1817, όταν η χολέρα επικεντρώθηκε μόνο στην Ανατολική και Νότια Ασία, κυρίως στην Ινδία, και δεν εξαπλώθηκε πέρα από τα σύνορά της.
II περίοδος - από το 1817 έως το 1926. Με την καθιέρωση ευρέων οικονομικών και άλλων δεσμών μεταξύ Ινδίας και ευρωπαϊκών και άλλων χωρών, η χολέρα ξεπέρασε την Ινδία και, εξαπλούμενη κατά μήκος των οδών των οικονομικών και θρησκευτικών δεσμών, προκάλεσε 6 πανδημίες που στοίχισαν εκατομμύρια ανθρώπινες ζωές. Η Ρωσία ήταν η πρώτη από τις ευρωπαϊκές χώρες όπου διείσδυσε η χολέρα. Από το 1823 έως το 1926, η Ρωσία βίωσε 57 χρόνια χολέρας. Κατά τη διάρκεια αυτής της περιόδου, περισσότεροι από 5,6 εκατομμύρια άνθρωποι αρρώστησαν με χολέρα και 2,14 εκατομμύρια άνθρωποι πέθαναν από αυτήν ("40%).
Περίοδος III - από το 1926 έως το 1961 Η χολέρα επέστρεψε στην κύρια ενδημική της εστία και ξεκίνησε μια περίοδος σχετικής ευημερίας. Φαινόταν ότι με την ανάπτυξη σύγχρονων συστημάτων καθαρισμού πόσιμου νερού, απομάκρυνσης και απολύμανσης λυμάτων και ανάπτυξης ειδικών μέτρων κατά της χολέρας, συμπεριλαμβανομένης της δημιουργίας υπηρεσίας καραντίνας, οι χώρες του κόσμου θα προστατεύονταν αξιόπιστα από μια άλλη εισβολή χολέρας.
Η τέταρτη περίοδος ξεκίνησε το 1961 και συνεχίζεται μέχρι σήμερα. Η έβδομη πανδημία δεν ξεκίνησε στην Ινδία, αλλά στην Ινδονησία, εξαπλώθηκε γρήγορα στις Φιλιππίνες, την Κίνα, τις χώρες της Ινδοκίνας και στη συνέχεια σε άλλες χώρες της Ασίας, της Αφρικής και της Ευρώπης. Οι ιδιαιτερότητες αυτής της πανδημίας περιλαμβάνουν το γεγονός ότι, πρώτον, προκλήθηκε από μια ειδική παραλλαγή του δονητικού ιού της χολέρας - V. cholerae eltor, η οποία μέχρι το 1961 δεν είχε καν αναγνωριστεί επίσημα ως ο αιτιολογικός παράγοντας της χολέρας. δεύτερον, όσον αφορά τη διάρκεια, ξεπέρασε όλες τις προηγούμενες πανδημίες. τρίτον, εμφανίστηκε σε δύο κύματα, το πρώτο από τα οποία διήρκεσε μέχρι το 1990 και το δεύτερο ξεκίνησε το 1991 και κάλυψε πολλές χώρες στη Νότια και Βόρεια Αμερική, συμπεριλαμβανομένων των Ηνωμένων Πολιτειών, οι οποίες δεν είχαν δει επιδημία χολέρας από το 1866. Από το 1961 έως το 1996, 3.943.239 άνθρωποι αρρώστησαν με χολέρα σε 146 χώρες.
Ο αιτιολογικός παράγοντας της χολέρας, το Vibrio cholerae, ανακαλύφθηκε το 1883 κατά τη διάρκεια της πέμπτης πανδημίας από τον R. Koch, αλλά το vibrio ανακαλύφθηκε για πρώτη φορά στα κόπρανα ασθενών με διάρροια το 1854 από τον F. Pacini.
Το V. cholerae ανήκει στην οικογένεια Vibrionaceae, η οποία περιλαμβάνει διάφορα γένη (Vibrio, Aeromonas, Plesiomonas, Photobacterium). Το γένος Vibrio έχει περισσότερα από 25 είδη από το 1985, εκ των οποίων τα πιο σημαντικά για τον άνθρωπο είναι τα V. cholerae, V. parahaemolyticus, V. alginolyticus, V. vulnificus και V. fluvialis.
Βασικά χαρακτηριστικά του γένους Vibrio
Κοντές, μη σχηματίζουσες σπόρια και κάψουλες, καμπύλες ή ευθείες αρνητικές κατά Gram ράβδοι, διαμέτρου 0,5 µm και μήκους 1,5-3,0 µm, κινητικές (το V. cholerae είναι μονότριχο, ορισμένα είδη έχουν δύο ή περισσότερα πολικά μαστίγια). Αναπτύσσονται καλά και γρήγορα σε κανονικά μέσα, είναι χημειοοργανοτρόφα και ζυμώνουν υδατάνθρακες για να παράγουν οξύ χωρίς αέριο (η γλυκόζη ζυμώνεται μέσω της οδού Embden-Meyerhof). Θετικές στην οξειδάση, σχηματίζουν ινδόλη, ανάγουν τα νιτρικά σε νιτρώδη (το V. cholerae δίνει θετική αντίδραση νιτροσοϊνδόλης), διασπούν τη ζελατίνη, συχνά δίνουν θετική αντίδραση Voges-Proskauer (δηλαδή, σχηματίζουν ακετυλομεθυλοκαρβινόλη), δεν έχουν ουρεάση, δεν σχηματίζουν H2S, έχουν λυσίνη και ορνιθίνη δεκαρβοξυλάσες, αλλά δεν έχουν αργινίνη διυδρολάση. Χαρακτηριστικό γνώρισμα του γένους Vibrio είναι η ευαισθησία των περισσότερων βακτηριακών στελεχών στο φάρμακο 0/129 (2,4-διαμινο-6,7-διαζοπροπυλπτεριδίνη), ενώ εκπρόσωποι των οικογενειών Pseudomonadaceae και Enterobacteriaceae είναι ανθεκτικοί σε αυτό το φάρμακο. Τα Vibrio είναι αερόβια και προαιρετικά αναερόβια, η βέλτιστη θερμοκρασία για ανάπτυξη είναι 18-37 C, pH 8,6-9,0 (αναπτύσσονται στην περιοχή pH 6,0-9,6), ορισμένα είδη (αλόφιλα) δεν αναπτύσσονται απουσία NaCl. Η περιεκτικότητα σε G + C στο DNA είναι 40-50 mol % (για το V. cholerae περίπου 47 mol %). Οι βιοχημικές δοκιμές χρησιμοποιούνται για τη διαφοροποίηση εντός της οικογένειας Vibrionaceae από τα μορφολογικά παρόμοια γένη Aeromonas και Plesiomonas, καθώς και για τη διάκριση από την οικογένεια Enterobacteriaceae.
Το δονάκιο χολέρας διαφέρει από την οικογένεια Pseudomonadaceae στο ότι ζυμώνει γλυκόζη μόνο μέσω της οδού Embden-Meyerhof (χωρίς τη συμμετοχή O2), ενώ τα πρώτα καταναλώνουν γλυκόζη μόνο παρουσία O2. Αυτή η διαφορά μεταξύ τους αποκαλύπτεται εύκολα στο μέσο Hugh-Leifson. Το μέσο περιέχει θρεπτικό άγαρ, γλυκόζη και έναν δείκτη. Η σπορά γίνεται σε δύο στήλες με το μέσο Hugh-Leifson, η μία εκ των οποίων είναι γεμάτη με βαζελίνη (για τη δημιουργία αναερόβιων συνθηκών). Στην περίπτωση ανάπτυξης του δονάκιου χολέρας, το χρώμα του μέσου αλλάζει και στους δύο δοκιμαστικούς σωλήνες, στην περίπτωση ανάπτυξης ψευδομονάδων - μόνο στον δοκιμαστικό σωλήνα χωρίς βαζελίνη (αερόβιες συνθήκες ανάπτυξης).
Το δονάκιο χολέρας δεν έχει πολλές απαιτήσεις σε θρεπτικά μέσα. Αναπαράγεται καλά και γρήγορα σε 1% αλκαλικό (pH 8,6-9,0) νερό πεπτόνης (PV) που περιέχει 0,5-1,0% NaCl, ξεπερνώντας την ανάπτυξη άλλων βακτηρίων. Για την καταστολή της ανάπτυξης του Proteus, συνιστάται η προσθήκη τελουρίτη καλίου (σε τελική αραίωση 1:100.000) σε 1% PV. Το 1% PV είναι το καλύτερο μέσο εμπλουτισμού για το δονάκιο χολέρας. Κατά τη διάρκεια της ανάπτυξης, σχηματίζει μια λεπτή, χαλαρή, γκριζωπή μεμβράνη στην επιφάνεια του PV μετά από 6-8 ώρες, η οποία καταστρέφεται εύκολα όταν ανακινείται και πέφτει στον πυθμένα με τη μορφή νιφάδων, το PV γίνεται μέτρια θολό. Έχουν προταθεί διάφορα επιλεκτικά μέσα για την απομόνωση του δονάκιου χολέρας: αλκαλικό άγαρ, άγαρ χολικού άλατος, αλκαλικό αλβουμινικό, αλκαλικό άγαρ με αίμα, λακτόζη-σακχαρόζη και άλλα μέσα. Το καλύτερο είναι το μέσο TCBS (άγαρ θειοθειικού κιτρικού-βρωμοθυμόλης σακχαρόζης) και οι τροποποιήσεις του. Ωστόσο, συχνότερα χρησιμοποιείται αλκαλικό MPA, στο οποίο το δονάκιο της χολέρας σχηματίζει λείες, υαλώδη-διαφανείς, μπλε-απόχρωσης, δισκοειδείς αποικίες με ιξώδη υφή.
Όταν σπέρνεται με έγχυση σε στήλη ζελατίνης, το δονάκιο, μετά από 2 ημέρες σε θερμοκρασία 22-23°C, προκαλεί υγροποίηση από την επιφάνεια με τη μορφή φυσαλίδας, στη συνέχεια χωνοειδούς και τέλος, στρώσης-στρώσης.
Στο γάλα, το δονητικό πολλαπλασιάζεται γρήγορα, προκαλώντας πήξη μετά από 24-48 ώρες, και στη συνέχεια συμβαίνει πεπτονίωση του γάλακτος, και μετά από 3-4 ημέρες το δονητικό πεθαίνει λόγω μετατόπισης του pH του γάλακτος προς την όξινη πλευρά.
Ο B. Heiberg, με βάση την ικανότητά τους να ζυμώνουν μαννόζη, σακχαρόζη και αραβινόζη, διαίρεσε όλα τα δονητικά (χολέρα και χολεροειδή) σε έναν αριθμό ομάδων, ο αριθμός των οποίων τώρα ανέρχεται σε 8.
Το Vibrio cholerae ανήκει στην πρώτη ομάδα του Heiberg.
Δονητικά στελέχη (Vibria) παρόμοια σε μορφολογικά, πολιτισμικά και βιοχημικά χαρακτηριστικά με το δονητικό στέλεχος της χολέρας ονομάζονταν και ονομάζονται διαφορετικά: παραχολέρα, χολεροειδή, NAG-δονητικά στελέχη (μη συγκολλητικά δονητικά στελέχη). Δονητικά στελέχη που δεν ανήκουν στην ομάδα O1. Το τελευταίο όνομα τονίζει με μεγαλύτερη ακρίβεια τη σχέση τους με το δονητικό στέλεχος της χολέρας. Όπως διαπιστώθηκε από τους A. Gardner και K. Venkat-Raman, τα δονητικά στελέχη της χολέρας και τα δονητικά στελέχη που μοιάζουν με χολέρα έχουν ένα κοινό Η-αντιγόνο, αλλά διαφέρουν στα Ο-αντιγόνα. Σύμφωνα με το Ο-αντιγόνο, τα δονητικά στελέχη της χολέρας και τα δονητικά στελέχη που μοιάζουν με χολέρα διαιρούνται επί του παρόντος σε 139 Ο-οροομάδες, αλλά ο αριθμός τους συνεχώς αυξάνεται. Το δονητικό στέλεχος της χολέρας ανήκει στην ομάδα O1. Έχει ένα κοινό Α-αντιγόνο και δύο αντιγόνα ειδικά για τον τύπο - Β και C, με τα οποία διακρίνονται τρεις ορότυποι του V. cholerae - ο ορότυπος Ogawa (AB), ο ορότυπος Inaba (AC) και ο ορότυπος Hikoshima (ABC). Το δονάκιο χολέρας στο στάδιο της διάσπασης έχει ένα αντιγόνο OR. Από αυτή την άποψη, ο ορός Ο, ο ορός OR και οι τυποειδικοί οροί Inaba και Ogawa χρησιμοποιούνται για την ταυτοποίηση του V. cholerae.
Το 1992-1993, ξεκίνησε μια μεγάλη επιδημία χολέρας στο Μπαγκλαντές, την Ινδία, την Κίνα, τη Μαλαισία και άλλες χώρες, ο αιτιολογικός παράγοντας της οποίας ήταν ένας νέος, προηγουμένως άγνωστος ορότυπος του είδους Vibrio cholerae. Διαφέρει από το V. cholerae O1 σε αντιγονικά χαρακτηριστικά: έχει το αντιγόνο 0139 και μια κάψουλα πολυσακχαρίτη και δεν συγκολλάται από κανέναν άλλο Ο-ορό. Όλες οι άλλες μορφολογικές και βιολογικές του ιδιότητες, συμπεριλαμβανομένης της ικανότητας να προκαλεί χολέρα, δηλαδή να συνθέτει την εξωτοξίνη-χολερογόνο, αποδείχθηκαν παρόμοιες με τις ιδιότητες του V. cholerae O1. Συνεπώς, ένας νέος αιτιολογικός παράγοντας της χολέρας, το V. cholerae 0139, προφανώς προέκυψε ως αποτέλεσμα μιας μετάλλαξης που άλλαξε το Ο-αντιγόνο. Ονομάστηκε V. cholerae 0139 bengal.
Το ζήτημα της σχέσης των λεγόμενων δονητών που μοιάζουν με χολέρα με το V. cholerae είναι από καιρό ασαφές. Ωστόσο, μια σύγκριση του V. cholerae και των δονητών που μοιάζουν με χολέρα (NAG-δονητών) με περισσότερα από 70 χαρακτηριστικά αποκάλυψε ομοιότητά τους κατά 90% και ο βαθμός ομολογίας DNA του V. cholerae και των μελετημένων NAG-δονητών είναι 70-100%. Επομένως, οι δονητές που μοιάζουν με χολέρα συνδυάζονται σε ένα είδος με το δονητή χολέρας, από το οποίο διαφέρουν κυρίως στα Ο-αντιγόνα τους, σε σχέση με τα οποία ονομάζονται δονητές της μη-01-ομάδας - V. cholerae μη-01.
Το είδος V. cholerae διαιρείται σε 4 βιότυπους: V. cholerae, V. eltor, V. proteus και V. albensis. Η φύση του δονητικού El Tor έχει συζητηθεί εδώ και πολλά χρόνια. Αυτό το δονητικό απομονώθηκε το 1906 από τον F. Gottschlich στον σταθμό καραντίνας El Tor από το σώμα ενός προσκυνητή που πέθανε από δυσεντερία. Ο F. Gottschlich απομόνωσε πολλά τέτοια στελέχη. Δεν διέφεραν από το δονητικό χολέρας σε όλες τις ιδιότητές τους και συγκολλήθηκαν με ορό χολέρας Ο. Ωστόσο, επειδή δεν υπήρχε χολέρα μεταξύ των προσκυνητών εκείνη την εποχή, και η μακροχρόνια μεταφορά του δονητικού χολέρας θεωρούνταν απίθανη, το ζήτημα του πιθανού αιτιολογικού ρόλου του V. eltor στη χολέρα παρέμεινε αμφιλεγόμενο για μεγάλο χρονικό διάστημα. Επιπλέον, το δονητικό El Tor, σε αντίθεση με το V. cholerae, είχε αιμολυτική δράση. Ωστόσο, το 1937, αυτό το δονητικό βακτήριο προκάλεσε μια μεγάλη και σοβαρή επιδημία χολέρας στο νησί Σουλαουέζι (Ινδονησία) με ποσοστό θνησιμότητας άνω του 60%. Τελικά, το 1961, έγινε ο ένοχος της 7ης πανδημίας και το 1962 το ζήτημα της χολερικής του φύσης επιλύθηκε οριστικά. Οι διαφορές μεταξύ του V. cholerae και του V. eltor αφορούν μόνο ορισμένα χαρακτηριστικά. Σε όλες τις άλλες ιδιότητες, το V. eltor δεν διαφέρει ουσιαστικά από το V. cholerae. Επιπλέον, έχει πλέον διαπιστωθεί ότι ο βιότυπος V. proteus (V.finklerpriori) περιλαμβάνει ολόκληρη την ομάδα των δονητών, εκτός από την ομάδα 01 (και τώρα 0139), που προηγουμένως ονομαζόταν δονητές NAG. Ο βιότυπος V. albensis απομονώθηκε από τον ποταμό Έλβα και έχει την ικανότητα να φωσφορίζει, αλλά έχοντας χάσει την ικανότητα αυτή, δεν διαφέρει από το V. proteus. Με βάση αυτά τα δεδομένα, το είδος Vibrio cholerae διαιρείται σήμερα σε 4 βιότυπους: V. cholerae 01 cholerae, V. cholerae eltor, V. cholerae 0139 bengal και V. cholerae non 01. Οι τρεις πρώτοι ανήκουν σε δύο ορότυπους 01 και 0139. Ο τελευταίος βιότυπος περιλαμβάνει τους προηγούμενους βιότυπους V. proteus και V. albensis και αντιπροσωπεύεται από πολλούς άλλους ορότυπους δονητών που δεν συγκολλούνται από ορούς 01 και 0139, δηλαδή δονητές NAG.
Παράγοντες παθογένειας του δονακίου χολέρας
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ]
Χημειοταξία του Vibrio cholerae
Με τη βοήθεια αυτών των ιδιοτήτων, το δονητικό αλληλεπιδρά με τα επιθηλιακά κύτταρα. Στα μεταλλαγμένα δονητικά χολερικά (που έχουν χάσει την ικανότητα χημειοταξίας), η λοιμογόνος δράση μειώνεται σημαντικά, ενώ στα μεταλλαγμένα Mob (που έχουν χάσει την κινητικότητά τους) είτε εξαφανίζεται εντελώς είτε μειώνεται απότομα.
Παράγοντες προσκόλλησης και αποικισμού μέσω των οποίων το δονάκιο προσκολλάται στις μικρολάχνες και αποικίζει την βλεννογόνο μεμβράνη του λεπτού εντέρου. Οι παράγοντες προσκόλλησης περιλαμβάνουν τη μουκινάση, τη διαλυτή αιμαγλουτινίνη/πρωτεάση, τη νευραμινιδάση κ.λπ. Προάγουν την προσκόλληση και τον αποικισμό καταστρέφοντας ουσίες που αποτελούν μέρος της βλέννας. Η διαλυτή αιμαγλουτινίνη/πρωτεάση προάγει τον διαχωρισμό των δονακίων από τους υποδοχείς των επιθηλιακών κυττάρων και την έξοδό τους από το έντερο στο εξωτερικό περιβάλλον, εξασφαλίζοντας την επιδημική τους εξάπλωση. Η νευραμινιδάση ενισχύει τον δεσμό μεταξύ του χολεραγόνου και των επιθηλιακών κυττάρων και διευκολύνει τη διείσδυση της τοξίνης στα κύτταρα, γεγονός που αυξάνει τη σοβαρότητα της διάρροιας.
Η τοξίνη της χολέρας είναι ένα χολερογόνο.
Οι λεγόμενες νέες τοξίνες που είναι ικανές να προκαλέσουν διάρροια, αλλά δεν έχουν γενετική ή ανοσολογική σχέση με τη χολεραγόνο.
Δερμονευρωτικοί και αιμορραγικοί παράγοντες. Η φύση αυτών των τοξικών παραγόντων και ο ρόλος τους στην παθογένεση της χολέρας δεν έχουν μελετηθεί επαρκώς.
[ 13 ], [ 14 ], [ 15 ], [ 16 ], [ 17 ], [ 18 ]
Ενδοτοξίνες του Vibrio cholerae
Οι λιποπολυσακχαρίτες του V. cholerae έχουν ισχυρή ενδοτοξική ιδιότητα και προκαλούν γενική δηλητηρίαση του σώματος.
Ο κύριος από τους αναφερόμενους παράγοντες παθογένειας του δονακίου χολέρας είναι η εξωτοξίνη χολέρα (CTX AB), η οποία καθορίζει την παθογένεση αυτής της ασθένειας. Το μόριο χολέρας αποτελείται από δύο θραύσματα - Α και Β. Το θραύσμα Α αποτελείται από δύο πεπτίδια - Α1 και Α2, έχει μια ειδική ιδιότητα τοξίνης χολέρας και της δίνει τις ιδιότητες ενός υπεραντιγόνου. Το θραύσμα Β αποτελείται από 5 πανομοιότυπες υπομονάδες. Εκτελεί δύο λειτουργίες: 1) αναγνωρίζει τον υποδοχέα (μονοσιαλογαγγλιοζίτη) του εντεροκυττάρου και συνδέεται με αυτόν· 2) σχηματίζει ένα ενδομεμβρανικό υδρόφοβο κανάλι για τη διέλευση της υπομονάδας Α. Το πεπτίδιο Α2 χρησιμεύει για τη σύνδεση των θραυσμάτων Α και Β. Η πραγματική τοξική λειτουργία εκτελείται από το πεπτίδιο Aj (ADP-ριβοζυλτρανσφεράση). Αλληλεπιδρά με το NAD, προκαλώντας την υδρόλυσή του· η προκύπτουσα ADP-ριβόζη συνδέεται με την ρυθμιστική υπομονάδα της αδενυλικής κυκλάσης. Αυτό οδηγεί σε αναστολή της υδρόλυσης GTP. Το προκύπτον σύμπλοκο GTP + αδενυλικής κυκλάσης προκαλεί υδρόλυση ATP με σχηματισμό cAMP. (Μια άλλη οδός για τη συσσώρευση cAMP είναι η καταστολή από το χολεραγόνο του ενζύμου που υδρολύει το cAMP σε 5-AMP). Η εκδήλωση της λειτουργίας του γονιδίου ctxAB που κωδικοποιεί τη σύνθεση της εξωτοξίνης εξαρτάται από τη λειτουργία ορισμένων άλλων γονιδίων παθογένειας, ιδιαίτερα των γονιδίων tcp (που κωδικοποιούν τη σύνθεση των ελεγχόμενων από την τοξίνη ινών προσκόλλησης - TCAP), των ρυθμιστικών γονιδίων toxR, toxS και toxT, των γονιδίων hap (διαλυτή αιμοσυγκολλητίνη/πρωτεάση) και νευραμινιδάσης (νευραμινιδάση). Επομένως, ο γενετικός έλεγχος της παθογένειας του V. cholerae είναι πολύπλοκος.
Όπως αποδείχθηκε, υπάρχουν δύο νησίδες παθογένειας στο χρωμόσωμα V. cholerae. Η μία από αυτές είναι το γονιδίωμα του νηματοειδούς μέτριου μετατρεπτικού φάγου CTXφ, και η άλλη είναι το γονιδίωμα του επίσης νηματοειδούς μέτριου μετατρεπτικού φάγου VPIcp. Κάθε μία από αυτές τις νησίδες παθογένειας περιέχει κασέτες γονιδίων που καθορίζονται στην πρόφαση, οι οποίες καθορίζουν την παθογένεια του παθογόνου της χολέρας. Το προφάγο CTXφ φέρει τα γονίδια CTX, γονίδια νέων τοξινών zot και ace, το γονίδιο ser (σύνθεση προσκολλητίνης) και το γονίδιο ortU (σύνθεση ενός προϊόντος με άγνωστη λειτουργία). Αυτή η κασέτα περιλαμβάνει επίσης το γονίδιο nei και την περιοχή φάγου RS2, η οποία κωδικοποιεί την αντιγραφή και την ενσωμάτωση του προφάγου στα χρωμοσώματα. Τα γονίδια zot, ace και ortU είναι απαραίτητα για τον σχηματισμό ιικών φάγων όταν το προφάγο αποκλείεται από το χρωμόσωμα του παθογόνου.
Το προφάγο VPIcp φέρει τα γονίδια tcp (που κωδικοποιούν την παραγωγή των τριχοειδών (πρωτεΐνη TCPA)), toxT, toxR, act (γονίδια πρόσθετου παράγοντα αποικισμού, κινητικότητας (ιντεγκράσες και τρανσποζάσες)). Η μεταγραφή των γονιδίων λοιμογόνου δράσης ρυθμίζεται από τρία ρυθμιστικά γονίδια: toxR, toxS και toxT. Αυτά τα γονίδια συντονισμένα, σε επίπεδο μεταγραφής, αλλάζουν τη δραστηριότητα περισσότερων από 20 γονιδίων λοιμογόνου δράσης, συμπεριλαμβανομένων των ctxAB, tcp και άλλων γονιδίων. Το κύριο ρυθμιστικό γονίδιο είναι το γονίδιο toxR. Η βλάβη ή η απουσία του οδηγεί σε μη λοιμογόνο δράση ή σε περισσότερο από 100 φορές μείωση στην παραγωγή της τοξίνης χολέρας CTX και TCPA. Πιθανώς, έτσι ρυθμίζεται η συντονισμένη έκφραση των γονιδίων λοιμογόνου δράσης σε νησίδες παθογένειας που σχηματίζονται από εύκρατους μετατρεπτικούς φάγους και σε άλλα βακτηριακά είδη. Έχει διαπιστωθεί ότι ένας άλλος προφάγος K139 υπάρχει στο χρωμόσωμα του V. cholerae eltor, αλλά το γονιδίωμά του έχει μελετηθεί ελάχιστα.
Το γονίδιο hap εντοπίζεται στο χρωμόσωμα. Έτσι, η λοιμογόνος δράση (παθογένεια) και η επιδημική ικανότητα του V. cholerae καθορίζονται από 4 γονίδια: ctxAB, tcp, toxR και hap.
Διάφορες μέθοδοι μπορούν να χρησιμοποιηθούν για την ανίχνευση της ικανότητας του V. cholerae να παράγει χολεραγόνο.
Βιολογική δοκιμή σε κουνέλια. Όταν τα δονάκια χολέρας εγχέονται ενδομυϊκά σε θηλάζοντα κουνέλια (ηλικίας όχι άνω των 2 εβδομάδων), αναπτύσσουν ένα τυπικό σύνδρομο χολέρας: διάρροια, αφυδάτωση και θάνατο του κουνελιού.
Άμεση ανίχνευση του χολερογόνου μέσω PCR, IFM ή παθητικής ανοσολογικής αντίδρασης αιμόλυσης (το χολερογόνο συνδέεται με την Gmj των ερυθροκυττάρων και αυτά λύονται με την προσθήκη αντιτοξικών αντισωμάτων και συμπληρώματος). Ωστόσο, η ανίχνευση της ικανότητας παραγωγής τοξίνης από μόνη της δεν επαρκεί για τον προσδιορισμό του επιδημικού κινδύνου τέτοιων στελεχών. Για αυτό, είναι απαραίτητο να ανιχνευθεί η παρουσία του γονιδίου hap, επομένως, ο καλύτερος και πιο αξιόπιστος τρόπος για τη διαφοροποίηση των τοξινογόνων και επιδημικών στελεχών των δονακίων χολέρας των οροομάδων 01 και 0139 είναι με PCR χρησιμοποιώντας ειδικούς εκκινητές για την ανίχνευση και των 4 γονιδίων παθογένειας: ctxAB, tcp, toxR και hap.
Η ικανότητα του V. cholerae, εκτός από τις οροομάδες 01 ή 0139, να προκαλεί σποραδικές ή αθροιστικές διαρροϊκές ασθένειες στους ανθρώπους μπορεί να οφείλεται είτε στην παρουσία εντεροτοξινών τύπου LT ή ST, οι οποίες διεγείρουν τα συστήματα αδενυλικής ή γουανυλικής κυκλάσης, αντίστοιχα, είτε στην παρουσία μόνο των γονιδίων ctxAB αλλά όχι του γονιδίου hap.
Κατά τη διάρκεια της έβδομης πανδημίας, απομονώθηκαν στελέχη V. cholerae με ποικίλους βαθμούς λοιμογόνου δράσης: χολερογόνα (λοιμογόνα), ασθενώς χολερογόνα (χαμηλής λοιμογόνου δράσης) και μη χολερογόνα (μη λοιμογόνα). Τα μη χολερογόνα V. cholerae, κατά κανόνα, εμφανίζουν αιμολυτική δράση, δεν λύονται από τον διαγνωστικό φάγο χολέρας HDF(5) και δεν προκαλούν ασθένειες στον άνθρωπο.
Για την τυποποίηση φάγων του V. cholerae 01 (συμπεριλαμβανομένου του El Tor), ο S. Mukherjee πρότεινε σύνολα φάγων, τα οποία στη συνέχεια συμπληρώθηκαν με άλλους φάγους στη Ρωσία. Ένα σύνολο τέτοιων φάγων (1-7) επιτρέπει τη διάκριση τύπων φάγων μεταξύ του V. cholerae 0116. Για την ταυτοποίηση του τοξικογόνου και μη τοξικογόνου V. cholerae El Tor, αντί των HDF-3, HDF-4 και HDF-5, προτείνονται πλέον στη Ρωσία οι φάγοι CTX* (λύουν τοξικογόνα δονάκια El Tor) και CTX" (λύουν μη τοξικογόνα δονάκια El Tor).
Ανθεκτικότητα σε παθογόνα χολέρας
Τα δονάκια χολέρας επιβιώνουν καλά σε χαμηλές θερμοκρασίες. Παραμένουν βιώσιμα στον πάγο για έως και 1 μήνα. Σε θαλασσινό νερό - έως και 47 ημέρες, σε ποτάμιο νερό - από 3-5 ημέρες έως αρκετές εβδομάδες, σε βραστό μεταλλικό νερό επιβιώνουν για περισσότερο από 1 χρόνο, στο έδαφος - από 8 ημέρες έως 3 μήνες, σε φρέσκα κόπρανα - έως και 3 ημέρες, σε βραστά προϊόντα (ρύζι, noodles, κρέας, χυλό κ.λπ.) επιβιώνουν για 2-5 ημέρες, σε ωμά λαχανικά - 2-4 ημέρες, σε φρούτα - 1-2 ημέρες, σε γάλα και γαλακτοκομικά προϊόντα - 5 ημέρες. Όταν φυλάσσονται στο κρύο, η περίοδος επιβίωσης αυξάνεται κατά 1-3 ημέρες. Σε λινό ύφασμα μολυσμένο με κόπρανα, επιβιώνουν έως και 2 ημέρες και σε υγρό υλικό - μια εβδομάδα. Τα δονάκια χολέρας πεθαίνουν μέσα σε 5 λεπτά σε θερμοκρασία 80 °C και ακαριαία στους 100 °C. Είναι ιδιαίτερα ευαίσθητα στα οξέα. Πεθαίνουν μέσα σε 5-15 λεπτά υπό την επίδραση χλωραμίνης και άλλων απολυμαντικών. Είναι ευαίσθητα στην ξήρανση και το άμεσο ηλιακό φως, αλλά επιβιώνουν καλά και για μεγάλο χρονικό διάστημα και πολλαπλασιάζονται ακόμη και σε ανοιχτά υδάτινα σώματα και λύματα πλούσια σε οργανική ύλη, με αλκαλικό pH και θερμοκρασία άνω των 10-12 °C. Είναι ιδιαίτερα ευαίσθητα στο χλώριο: μια δόση ενεργού χλωρίου 0,3-0,4 mg/l νερού σε 30 λεπτά προκαλεί αξιόπιστη απολύμανση από τα δονάκια χολέρας.
Δονάκια παθογόνα για τον άνθρωπο που δεν ανήκουν στο είδος Vibrio Cholerae
Το γένος Vibrio περιλαμβάνει περισσότερα από 25 είδη, από τα οποία, εκτός από το V. cholerae, τουλάχιστον οκτώ είναι ικανά να προκαλέσουν ασθένειες στους ανθρώπους: V. parahaemolyticus, V. alginolyticus, V. vulnificus, V. fluvialis, V. fumissii, V. mimicus, V. damsela και V. hollisae. Όλα αυτά τα δονητικά κατοικούν σε θάλασσες και κόλπους. Η μόλυνση γίνεται είτε μέσω της κολύμβησης είτε μέσω της κατανάλωσης θαλασσινών. Έχει διαπιστωθεί ότι τα δονητικά χολερικά και μη χολερικά μπορούν να προκαλέσουν όχι μόνο γαστρεντερίτιδα, αλλά και μολύνσεις τραυμάτων. Αυτή η ικανότητα έχει διαπιστωθεί στις ομάδες V. cholerae 01 και μη 01, V. parahaemolyticus, V. alginolyticus, V. mimicus, V. damsela και V. vulnificus. Προκαλούν φλεγμονώδεις διεργασίες στους μαλακούς ιστούς όταν υποβάλλονται σε βλάβη από το κέλυφος θαλάσσιων ζώων ή όταν έρχονται σε άμεση επαφή με μολυσμένο θαλασσινό νερό.
Από τα καταγεγραμμένα παθογόνα μη χολερικά δονητικά στελέχη, τα πιο πρακτικά ενδιαφέροντα είναι τα V. parahaemolyticus, V. alginolyticus, V. vulnificus και V. fluvialis.
Το V. parahaemolyticus - ένα παρααιμολυτικό δονητικό - απομονώθηκε για πρώτη φορά στην Ιαπωνία το 1950 κατά τη διάρκεια μιας μεγάλης επιδημίας τροφικής δηλητηρίασης που προκλήθηκε από την κατανάλωση ημιαποξηραμένων σαρδελών (θνησιμότητα 7,5%). Ο αιτιολογικός παράγοντας ανήκε στο γένος Vibrio από τον R. Sakazaki το 1963. Διαίρεσε τα μελετηθέντα στελέχη σε 2 είδη: V. parahaemolyticus και V. alginolyticus. Και τα δύο είδη βρίσκονται σε παράκτια θαλάσσια ύδατα και στους κατοίκους τους είναι αλόφιλα (ελληνικά hals - αλάτι). σε αντίθεση με τα συνηθισμένα δονητικά, τα αλόφιλα δεν αναπτύσσονται σε μέσα χωρίς NaCl και αναπαράγονται καλά σε υψηλές συγκεντρώσεις αυτού. Η σχέση των αλόφιλων δονικών καθορίζεται από την ικανότητά τους να ζυμώνουν σακχαρόζη, να σχηματίζουν ακετυλομεθυλοκαρβινόλη και να αναπαράγονται σε PV με 10% NaCl. Όλα αυτά τα χαρακτηριστικά είναι εγγενή στο είδος V. alginolyticus, αλλά απουσιάζουν από το V. parahaemolyticus.
Το παρααιμολυτικό δονητικό έχει τρεις τύπους αντιγόνων: θερμοευαίσθητα μαστιγωτά Η-αντιγόνα, θερμοσταθερά Ο-αντιγόνα που δεν καταστρέφονται με θέρμανση στους 120 °C για 2 ώρες και επιφανειακά Κ-αντιγόνα που καταστρέφονται με θέρμανση. Οι πρόσφατα απομονωμένες καλλιέργειες του V. parahaemolyticus έχουν σαφώς καθορισμένα Κ-αντιγόνα που προστατεύουν τα ζωντανά δονητικά από τη συγκόλληση από ομόλογους Ο-ορούς. Τα Η-αντιγόνα είναι τα ίδια για όλα τα στελέχη, αλλά τα Η-αντιγόνα του monotrichus διαφέρουν από τα Η-αντιγόνα των peritrichs. Σύμφωνα με το Ο-αντιγόνο, το V. parahaemolyticus διαιρείται σε 14 οροομάδες. Εντός των οροομάδων, τα δονητικά διαιρούνται σε ορότυπους σύμφωνα με τα Κ-αντιγόνα, ο συνολικός αριθμός των οποίων είναι 61. Το αντιγονικό σχήμα του V. parahaemolyticus έχει αναπτυχθεί μόνο για τα στελέχη του που απομονώθηκαν από ανθρώπους.
Η παθογένεση του V. parahaemolyticus σχετίζεται με την ικανότητά του να συνθέτει αιμολυσίνη, η οποία έχει εντεροτοξικές ιδιότητες. Η τελευταία ανιχνεύεται χρησιμοποιώντας τη μέθοδο Kanagawa. Η ουσία του έγκειται στο γεγονός ότι το V. parahaemolyticus, παθογόνο για τον άνθρωπο, προκαλεί σαφή αιμόλυση σε άγαρ αίματος που περιέχει 7% NaCl. Σε άγαρ αίματος που περιέχει λιγότερο από 5% NaCl, η αιμόλυση προκαλείται από πολλά στελέχη του V. parahaemolyticus, και σε άγαρ αίματος με 7% NaCl - μόνο στελέχη με εντεροπαθογόνες ιδιότητες. Το παρααιμολυτικό δονάκιο βρίσκεται στις ακτές της Ιαπωνικής, της Κασπίας, της Μαύρης και άλλων θαλασσών. Προκαλεί τροφιμογενείς τοξικές λοιμώξεις και ασθένειες που μοιάζουν με δυσεντερία. Η μόλυνση εμφανίζεται κατά την κατανάλωση ωμών ή ημιωμών θαλασσινών μολυσμένων με V-parahaemolyticus (θαλάσσια ψάρια, στρείδια, καρκινοειδή κ.λπ.).
Μεταξύ των οκτώ μη χολερικών δονητών που αναφέρθηκαν παραπάνω, το πιο παθογόνο για τον άνθρωπο είναι το V. vulnificus, το οποίο περιγράφηκε για πρώτη φορά το 1976 ως Beneckea vulnificus και στη συνέχεια αναταξινομήθηκε ως Vibrio vulnificus το 1980. Βρίσκεται συχνά στο θαλασσινό νερό και στους κατοίκους του και προκαλεί διάφορες ανθρώπινες ασθένειες. Τα στελέχη του V. vulnificus θαλάσσιας και κλινικής προέλευσης δεν διαφέρουν μεταξύ τους ούτε φαινοτυπικά ούτε γενετικά.
Οι λοιμώξεις τραυμάτων που προκαλούνται από το V. vulnificus εξελίσσονται ταχέως και οδηγούν στο σχηματισμό όγκων με επακόλουθη νέκρωση ιστών, που συνοδεύεται από πυρετό, ρίγη, μερικές φορές έντονο πόνο και, σε ορισμένες περιπτώσεις, απαιτεί ακρωτηριασμό.
Έχει διαπιστωθεί ότι το V. vulnificus παράγει εξωτοξίνη. Πειράματα σε ζώα έχουν δείξει ότι το παθογόνο προκαλεί σοβαρή τοπική βλάβη με την ανάπτυξη οιδήματος και νέκρωσης ιστών, που ακολουθείται από θάνατο. Ο ρόλος της εξωτοξίνης στην παθογένεση της νόσου βρίσκεται υπό μελέτη.
Εκτός από τις μολύνσεις τραυμάτων, το V. vulnificus μπορεί να προκαλέσει πνευμονία σε θύματα πνιγμού και ενδομητρίτιδα σε γυναίκες μετά από έκθεση σε θαλασσινό νερό. Η πιο σοβαρή μορφή λοίμωξης που προκαλείται από το V. vulnificus είναι η πρωτοπαθής σηψαιμία που σχετίζεται με την κατανάλωση ωμών στρειδιών (και πιθανώς άλλων θαλάσσιων ζώων). Αυτή η ασθένεια εξελίσσεται πολύ γρήγορα: ο ασθενής εμφανίζει αδιαθεσία, πυρετό, ρίγη και εξάντληση, και στη συνέχεια σοβαρή υπόταση, η οποία είναι η κύρια αιτία θανάτου (το ποσοστό θνησιμότητας είναι περίπου 50%).
Το V. fluvialis περιγράφηκε για πρώτη φορά ως παθογόνο γαστρεντερίτιδας το 1981. Ανήκει σε μια υποομάδα μη παθογόνων δονακίων χολέρας που έχουν διυδρολάση αργινίνης αλλά όχι νετορνιθίνη και λυσίνη δεκαρβοξυλάσες (V. fluvialis, V. furnissii, V. damsela, δηλαδή, φαινοτυπικά παρόμοια με την Aeromonas). Το V. fluvialis είναι ένας κοινός αιτιολογικός παράγοντας γαστρεντερίτιδας, η οποία συνοδεύεται από σοβαρό έμετο, διάρροια, κοιλιακό άλγος, πυρετό και σοβαρή ή μέτρια αφυδάτωση. Ο κύριος παθογόνος παράγοντας είναι η εντεροτοξίνη.
Επιδημιολογία της χολέρας
Η κύρια πηγή μόλυνσης είναι μόνο ένα άτομο - ένας ασθενής με χολέρα ή ένας φορέας δονητικού ιού, καθώς και το νερό που έχει μολυνθεί με αυτά. Κανένα ζώο στη φύση δεν μολύνεται από χολέρα. Η οδός μόλυνσης είναι η κοπρανοστοματική. Οδοί μόλυνσης: α) η κύρια - μέσω του νερού που χρησιμοποιείται για πόση, μπάνιο και οικιακές ανάγκες· β) επαφή-νοικοκυριό και γ) μέσω τροφής. Όλες οι μεγάλες επιδημίες και πανδημίες χολέρας συνδέονταν με το νερό. Τα δονητικά ιούς της χολέρας έχουν τέτοιους προσαρμοστικούς μηχανισμούς που διασφαλίζουν την ύπαρξη των πληθυσμών τους τόσο στο ανθρώπινο σώμα όσο και σε ορισμένα οικοσυστήματα ανοιχτών υδάτινων σωμάτων. Η σοβαρή διάρροια, η οποία προκαλείται από το δονητικό ιού της χολέρας, οδηγεί στον καθαρισμό των εντέρων από ανταγωνιστικά βακτήρια και συμβάλλει στην ευρεία εξάπλωση του παθογόνου στο περιβάλλον, κυρίως στα λύματα και σε ανοιχτά υδάτινα σώματα όπου απορρίπτονται. Ένα άτομο με χολέρα εκκρίνει το παθογόνο σε τεράστιες ποσότητες - από 100 εκατομμύρια έως 1 δισεκατομμύριο ανά 1 ml κοπράνων, ένας φορέας δονητικού ιού εκκρίνει 100-100.000 δονητικά ιούς σε 1 ml, η μολυσματική δόση είναι περίπου 1 εκατομμύριο δονητικά ιούς. Η διάρκεια απέκκρισης του χολερικού δονακίου σε υγιείς φορείς είναι από 7 έως 42 ημέρες και 7-10 ημέρες σε όσους έχουν αναρρώσει. Η μεγαλύτερη απέκκριση είναι εξαιρετικά σπάνια.
Μια ιδιαιτερότητα της χολέρας είναι ότι μετά από αυτήν, κατά κανόνα, δεν υπάρχει μακροχρόνια μεταφορά και δεν σχηματίζονται σταθερές ενδημικές εστίες. Ωστόσο, όπως ήδη αναφέρθηκε παραπάνω, λόγω της ρύπανσης των ανοιχτών υδάτινων σωμάτων με λύματα που περιέχουν μεγάλες ποσότητες οργανικών ουσιών, απορρυπαντικών και επιτραπέζιου αλατιού, το καλοκαίρι το δονητικό της χολέρας όχι μόνο επιβιώνει σε αυτά για μεγάλο χρονικό διάστημα, αλλά και πολλαπλασιάζεται.
Μεγάλης επιδημιολογικής σημασίας είναι το γεγονός ότι τα δονάκια χολέρας της ομάδας 01, τόσο τα μη τοξικογόνα όσο και τα τοξικογόνα, μπορούν να επιβιώσουν για μεγάλο χρονικό διάστημα σε διάφορα υδάτινα οικοσυστήματα ως ακαλλιέργητες μορφές. Χρησιμοποιώντας την αλυσιδωτή αντίδραση πολυμεράσης, ανιχνεύθηκαν γονίδια vct ακαλλιέργητων μορφών του V. chokrae σε διάφορα υδάτινα σώματα σε μια σειρά ενδημικών περιοχών της ΚΑΚ κατά τη διάρκεια αρνητικών βακτηριολογικών μελετών.
Η ενδημική εστία του δονητικού ιού χολέρας El Tor είναι η Ινδονησία, η εμφάνιση αυτού του ενόχου της έβδομης πανδημίας από εκεί πιστεύεται ότι σχετίζεται με την επέκταση των οικονομικών δεσμών της Ινδονησίας με τον έξω κόσμο μετά την ανεξαρτησία της, και η διάρκεια και η αστραπιαία ανάπτυξη της πανδημίας, ειδικά του δεύτερου κύματος της, επηρεάστηκαν αποφασιστικά από την έλλειψη ανοσίας στη χολέρα και διάφορες κοινωνικές αναταραχές στις χώρες της Ασίας, της Αφρικής και της Αμερικής.
Σε περίπτωση χολέρας, λαμβάνεται μια σειρά από αντιεπιδημικά μέτρα, μεταξύ των οποίων τα κορυφαία και αποφασιστικά είναι η ενεργός, έγκαιρη ανίχνευση και απομόνωση (νοσηλεία, θεραπεία) ασθενών σε οξείες και άτυπες μορφές και υγιών φορέων δονητικού ιού. Λαμβάνονται μέτρα για την πρόληψη πιθανών οδών μόλυνσης. Δίνεται ιδιαίτερη προσοχή στην παροχή νερού (χλωρίωση πόσιμου νερού), στη συμμόρφωση με τις υγειονομικές συνθήκες σε επιχειρήσεις τροφίμων, σε παιδικά ιδρύματα, σε δημόσιους χώρους. Πραγματοποιείται αυστηρός έλεγχος, συμπεριλαμβανομένου του βακτηριολογικού, σε ανοιχτά υδάτινα σώματα, πραγματοποιείται ανοσοποίηση του πληθυσμού κ.λπ.
Συμπτώματα χολέρας
Η περίοδος επώασης για τη χολέρα κυμαίνεται από μερικές ώρες έως 6 ημέρες, συνήθως 2-3 ημέρες. Αφού εισέλθουν στον αυλό του λεπτού εντέρου, τα δονάκια χολέρας, λόγω της κινητικότητάς τους και της χημειοταξίας τους προς τη βλεννογόνο μεμβράνη, κατευθύνονται προς τη βλέννα. Για να διεισδύσουν μέσα από αυτήν, τα δονάκια παράγουν μια σειρά από ένζυμα: νευραμινιδάση, μουκινάση, πρωτεάσες, λεκιθινάση, τα οποία καταστρέφουν τις ουσίες που περιέχονται στη βλέννα και διευκολύνουν την κίνηση των δονακίων στα επιθηλιακά κύτταρα. Με προσκόλληση, τα δονάκια προσκολλώνται στον γλυκοκάλυκα του επιθηλίου και, χάνοντας την κινητικότητά τους, αρχίζουν να πολλαπλασιάζονται εντατικά, αποικίζοντας τις μικρολάχνες του λεπτού εντέρου (βλ. ένθετο χρώματος, Εικ. 101.2) και ταυτόχρονα παράγουν μεγάλη ποσότητα εξωτοξίνης-χολερογόνου. Τα μόρια χολερογόνου συνδέονται με τη μονοσιαλογαγγλιοσίδη Gni! Και διεισδύουν στην κυτταρική μεμβράνη, όπου ενεργοποιούν το σύστημα αδενυλικής κυκλάσης, και η συσσωρευμένη cAMP προκαλεί υπερέκκριση υγρών, κατιόντων και ανιόντων Na, HCO3, Kl, Cl από τα εντεροκύτταρα, η οποία οδηγεί σε διάρροια χολέρας, αφυδάτωση και αφαλάτωση του σώματος. Υπάρχουν τρεις τύποι της νόσου:
- μια βίαιη, σοβαρή αφυδατωτική διάρροια που έχει ως αποτέλεσμα τον θάνατο του ασθενούς μέσα σε λίγες ώρες.
- λιγότερο σοβαρή πορεία ή διάρροια χωρίς αφυδάτωση.
- ασυμπτωματική πορεία της νόσου (μεταφορά δονητών).
Σε σοβαρές περιπτώσεις χολέρας, οι ασθενείς εμφανίζουν διάρροια, αυξάνεται η συχνότητα των κοπράνων, τα κόπρανα γίνονται πιο άφθονα, υδαρή, χάνουν την οσμή τους και μοιάζουν με ζωμό ρυζιού (ένα θολό υγρό με υπολείμματα βλέννας και επιθηλιακά κύτταρα που επιπλέουν σε αυτό). Στη συνέχεια εμφανίζεται εξουθενωτικός έμετος, πρώτα από το εντερικό περιεχόμενο, και στη συνέχεια ο έμετος παίρνει την εμφάνιση ζωμού ρυζιού. Η θερμοκρασία του ασθενούς πέφτει κάτω από το φυσιολογικό, το δέρμα γίνεται μπλε, ζαρωμένο και κρύο - χολέρα αλγικό. Ως αποτέλεσμα της αφυδάτωσης, το αίμα πήζει, αναπτύσσεται κυάνωση, η οξυγονωμένη λειτουργία, η νεφρική λειτουργία υποφέρει έντονα, εμφανίζονται σπασμοί, ο ασθενής χάνει τις αισθήσεις του και επέρχεται θάνατος. Το ποσοστό θνησιμότητας από χολέρα κατά τη διάρκεια της έβδομης πανδημίας κυμαινόταν από 1,5% στις ανεπτυγμένες χώρες έως 50% στις αναπτυσσόμενες χώρες.
Η μεταλοιμώδης ανοσία είναι ισχυρή, μακράς διαρκείας και οι υποτροπιάζουσες ασθένειες είναι σπάνιες. Η ανοσία είναι αντιτοξική και αντιμικροβιακή, και προκαλείται από αντισώματα (οι αντιτοξίνες παραμένουν για μεγαλύτερο χρονικό διάστημα από τα αντιμικροβιακά αντισώματα), κύτταρα ανοσολογικής μνήμης και φαγοκύτταρα.
Εργαστηριακή διάγνωση της χολέρας
Η κύρια και αποφασιστική μέθοδος διάγνωσης της χολέρας είναι η βακτηριολογική. Το υλικό για εξέταση από τον ασθενή είναι τα κόπρανα και ο εμετός. Τα κόπρανα εξετάζονται για τη μεταφορά δονητών. Ένα απολινωμένο τμήμα του λεπτού εντέρου και της χοληδόχου κύστης λαμβάνεται για εξέταση από άτομα που πέθαναν από χολέρα. Από αντικείμενα του εξωτερικού περιβάλλοντος, εξετάζεται συχνότερα νερό από ανοιχτές δεξαμενές και λύματα.
Κατά τη διεξαγωγή βακτηριολογικής μελέτης, πρέπει να πληρούνται οι ακόλουθες τρεις προϋποθέσεις:
- σπείρετε υλικό από τον ασθενή το συντομότερο δυνατό (το vibrio της χολέρας επιβιώνει στα κόπρανα για μικρό χρονικό διάστημα).
- το δοχείο στο οποίο λαμβάνεται το υλικό δεν πρέπει να απολυμαίνεται με χημικά και δεν πρέπει να περιέχει ίχνη αυτών, καθώς το δονητικό χολερικό είναι πολύ ευαίσθητο σε αυτά.
- να αποκλειστεί η πιθανότητα μόλυνσης και μόλυνσης άλλων.
Η καλλιέργεια απομονώνεται σύμφωνα με το ακόλουθο σχήμα: σπορά σε PV, ταυτόχρονα σε αλκαλικό MPA ή σε οποιοδήποτε επιλεκτικό μέσο (το TCBS είναι το καλύτερο). Μετά από 6 ώρες, εξετάζεται η μεμβράνη που σχηματίζεται στο PV και, εάν είναι απαραίτητο, γίνεται μεταφορά σε ένα δεύτερο PV (ο ρυθμός σποράς του δονητικού χολερικού σε αυτή την περίπτωση αυξάνεται κατά 10%). Από το PV, γίνεται μεταφορά σε αλκαλικό MPA. Οι ύποπτες αποικίες (υαλώδεις-διαφανείς) μεταφέρονται για να ληφθεί μια καθαρή καλλιέργεια, η οποία αναγνωρίζεται από μορφολογικές, καλλιεργητικές, βιοχημικές ιδιότητες, κινητικότητα και τελικά τυποποιείται χρησιμοποιώντας διαγνωστικούς ορούς συγκόλλησης O-, OR-, Inaba και Ogawa και φάγους (HDF). Έχουν προταθεί διάφορες επιλογές για επιταχυνόμενη διάγνωση, η καλύτερη από τις οποίες είναι η φωταυγής-ορολογική μέθοδος. Επιτρέπει την ανίχνευση δονητικού χολερικού απευθείας στο υλικό δοκιμής (ή μετά από προκαταρκτική καλλιέργεια σε δύο δοκιμαστικούς σωλήνες με 1% PV, στον έναν από τους οποίους προστίθεται ο φάγος χολέρας) εντός 1,5-2 ωρών. Για την επιταχυνόμενη ανίχνευση του δονακίου χολέρας, το IEM του Νίζνι Νόβγκοροντ πρότεινε ένα σύνολο δίσκων δείκτη χαρτιού που αποτελούνται από 13 βιοχημικές εξετάσεις (οξειδάση, ινδόλη, ουρεάση, λακτόζη, γλυκόζη, σακχαρόζη, μαννόζη, αραβινόζη, μαννιτόλη, ινοσιτόλη, αργινίνη, ορνιθίνη, λυσίνη), οι οποίες επιτρέπουν τη διαφοροποίηση εκπροσώπων του γένους Vibrio από τα γένη Aeromonas, Plesiomonas, Pseudomonas, Comamonas και από την οικογένεια Enterobacteriaceae. Για την ταχεία ανίχνευση του δονακίου χολέρας σε κόπρανα και σε περιβαλλοντικά αντικείμενα, μπορεί να χρησιμοποιηθεί RPGA με διαγνωστικό αντισωμάτων. Για την ανίχνευση μη καλλιεργημένων μορφών δονακίου χολέρας σε περιβαλλοντικά αντικείμενα, χρησιμοποιείται μόνο η μέθοδος αλυσιδωτής αντίδρασης πολυμεράσης.
Σε περιπτώσεις όπου απομονώνονται V. cholerae εκτός της ομάδας Ol, θα πρέπει να τυποποιούνται χρησιμοποιώντας τους αντίστοιχους ορούς συγκόλλησης άλλων οροομάδων. Η απομόνωση V. cholerae εκτός της ομάδας Ol από ασθενή με διάρροια (συμπεριλαμβανομένης της διάρροιας που μοιάζει με χολέρα) απαιτεί τα ίδια αντιεπιδημικά μέτρα όπως στην περίπτωση της απομόνωσης V. cholerae της ομάδας Ol. Εάν είναι απαραίτητο, η παρουσία των γονιδίων παθογένειας ctxAB, tcp, toxR και hap προσδιορίζεται σε τέτοια δονάκια χρησιμοποιώντας PCR.
Η ορολογική διάγνωση της χολέρας είναι βοηθητικής φύσης. Για το σκοπό αυτό, μπορεί να χρησιμοποιηθεί η αντίδραση συγκόλλησης, αλλά είναι καλύτερο να προσδιοριστεί ο τίτλος των δονικοκτόνων αντισωμάτων ή των αντιτοξινών (τα αντισώματα κατά της χολέρας προσδιορίζονται με ενζυμική ανοσοδοκιμασία ή μεθόδους ανοσοφθορισμού).
Εργαστηριακή διάγνωση μη παθογόνων δονακίων που προκαλούν χολέρα
Η κύρια μέθοδος διάγνωσης ασθενειών που προκαλούνται από μη παθογόνα δονάκια που δεν προκαλούν χολέρα είναι η βακτηριολογική χρησιμοποιώντας επιλεκτικά μέσα όπως TCBS, MacConkey, κ.λπ. Η ένταξη της απομονωμένης καλλιέργειας στο γένος Vibrio προσδιορίζεται με βάση τα βασικά χαρακτηριστικά των βακτηρίων αυτού του γένους.
Θεραπεία της χολέρας
Η θεραπεία των ασθενών με χολέρα θα πρέπει να συνίσταται κυρίως στην ενυδάτωση και την αποκατάσταση του φυσιολογικού μεταβολισμού νερού-αλατιού. Για το σκοπό αυτό, συνιστάται η χρήση αλατούχων διαλυμάτων, για παράδειγμα, με την ακόλουθη σύνθεση: NaCl - 3,5, NaHC03 - 2,5, KCl - 1,5 και γλυκόζη - 20,0 g ανά 1 λίτρο νερού. Αυτή η παθογενετικά τεκμηριωμένη θεραπεία σε συνδυασμό με ορθολογική αντιβιοτική θεραπεία επιτρέπει τη μείωση του ποσοστού θνησιμότητας στη χολέρα σε 1% ή λιγότερο.
Ειδική πρόληψη της χολέρας
Για τη δημιουργία τεχνητής ανοσίας, προτάθηκε ένα εμβόλιο χολέρας, συμπεριλαμβανομένου ενός που παρασκευάζεται από νεκρά στελέχη Inaba και Ogawa, ένα τοξοειδές χολέρας για υποδόρια χρήση και ένα εντερικό χημικό δισθενές εμβόλιο που αποτελείται από ανατοξίνη και σωματικά αντιγόνα των οροτύπων Inaba και Ogawa, καθώς δεν σχηματίζεται διασταυρούμενη ανοσία. Ωστόσο, η διάρκεια της ανοσίας μετά τον εμβολιασμό δεν υπερβαίνει τους 6-8 μήνες, επομένως οι εμβολιασμοί πραγματοποιούνται μόνο σύμφωνα με επιδημιολογικές ενδείξεις. Η προφύλαξη με αντιβιοτικά έχει αποδειχθεί καλή σε εστίες χολέρας, ιδιαίτερα στην τετρακυκλίνη, στην οποία το δονάκιο χολέρας είναι ιδιαίτερα ευαίσθητο. Άλλα αντιβιοτικά αποτελεσματικά κατά του V. cholerae μπορούν να χρησιμοποιηθούν για τον ίδιο σκοπό.